
Yphthimoides inornata ( Hayward, 1962 ), 2022
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5116.4.4 |
|
publication LSID |
lsid:zoobank.org:pub:036B84AB-790E-48E5-8657-B3D3DA143740 |
|
DOI |
https://doi.org/10.5281/zenodo.6377212 |
|
persistent identifier |
https://treatment.plazi.org/id/D5100520-FFEA-FFED-91EA-C59C9DBCB4C0 |
|
treatment provided by |
Plazi |
|
scientific name |
Yphthimoides inornata ( Hayward, 1962 ) |
| status |
stat. nov. |
Yphthimoides inornata ( Hayward, 1962) stat. rev.
( Figs 1–8 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 )
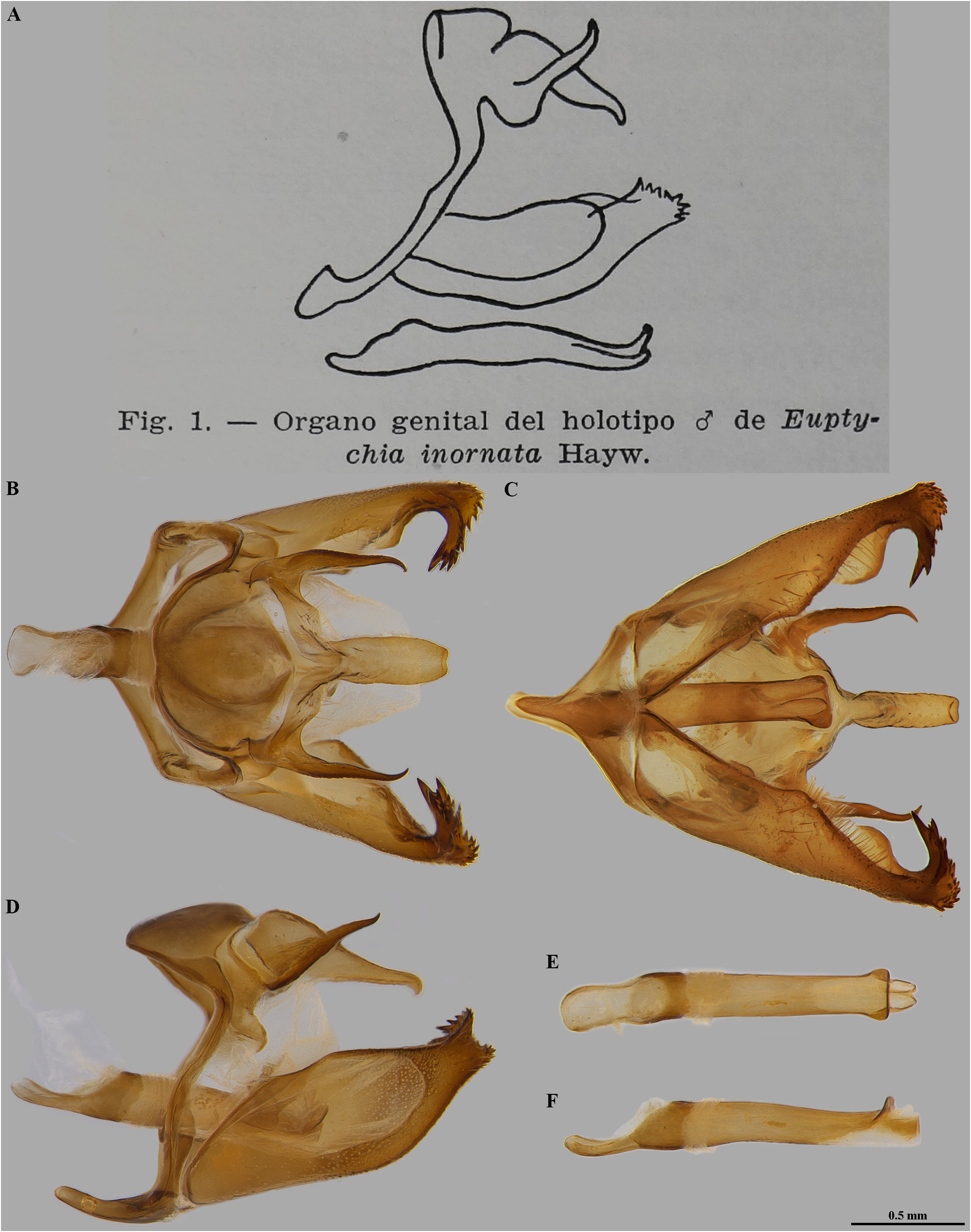
Euptychia inornata Hayward, 1962 . Acta zool. lill. 18: 12, fig. 1 (male genitalia).
Euptychia inornata was described based on the holotype male ( Fig. 1 View FIGURE 1 ) from Pelotas, Rio Grande do Sul, Brazil, Biezanko leg., XI-1953 and the allotype female from Playadito, Corrientes, Argentina, Gibson leg., X-1953, both in Instituto Miguel Lillo [Fundación Miguel Lillo, Tucumán, Argentina]; the holotype genitalia was illustrated.
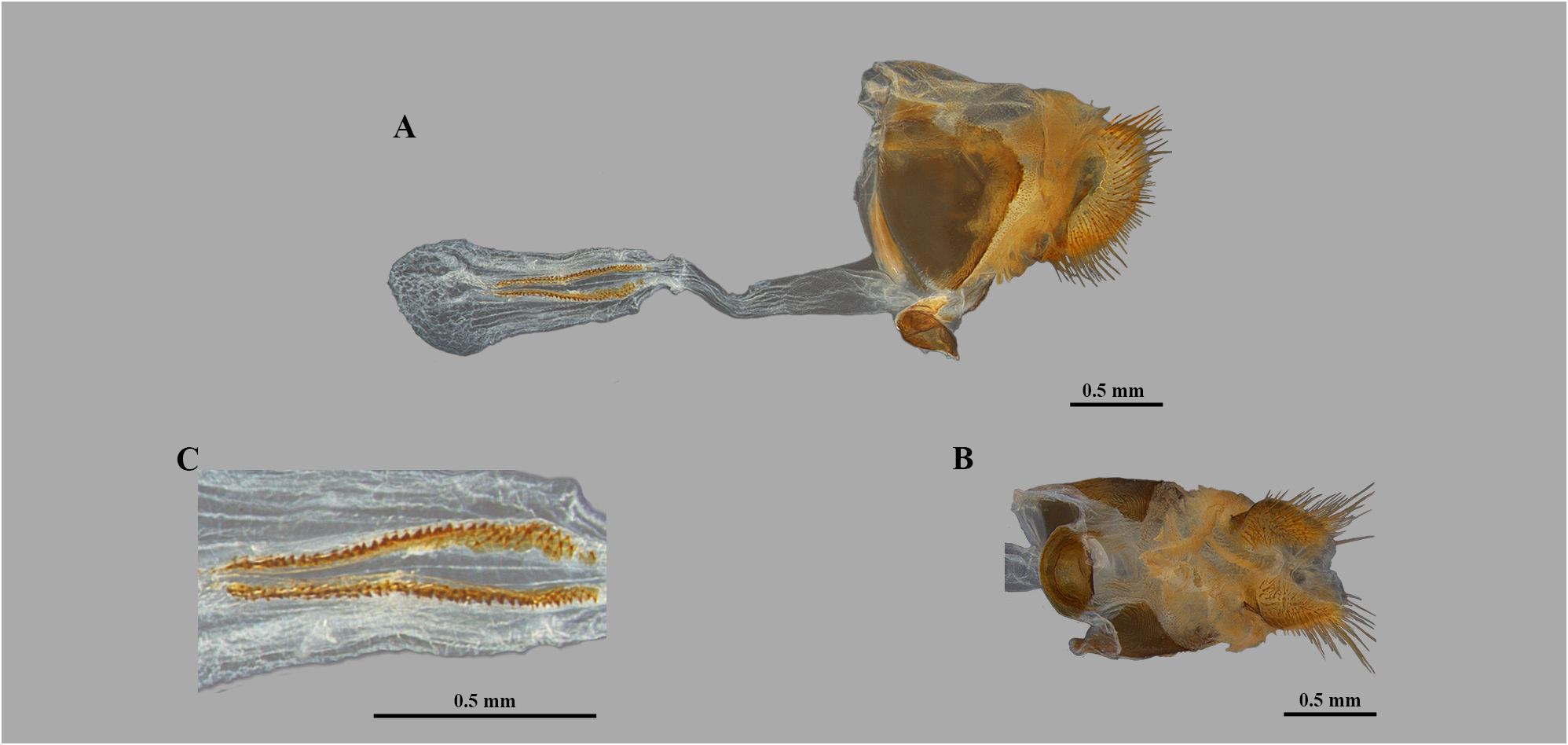
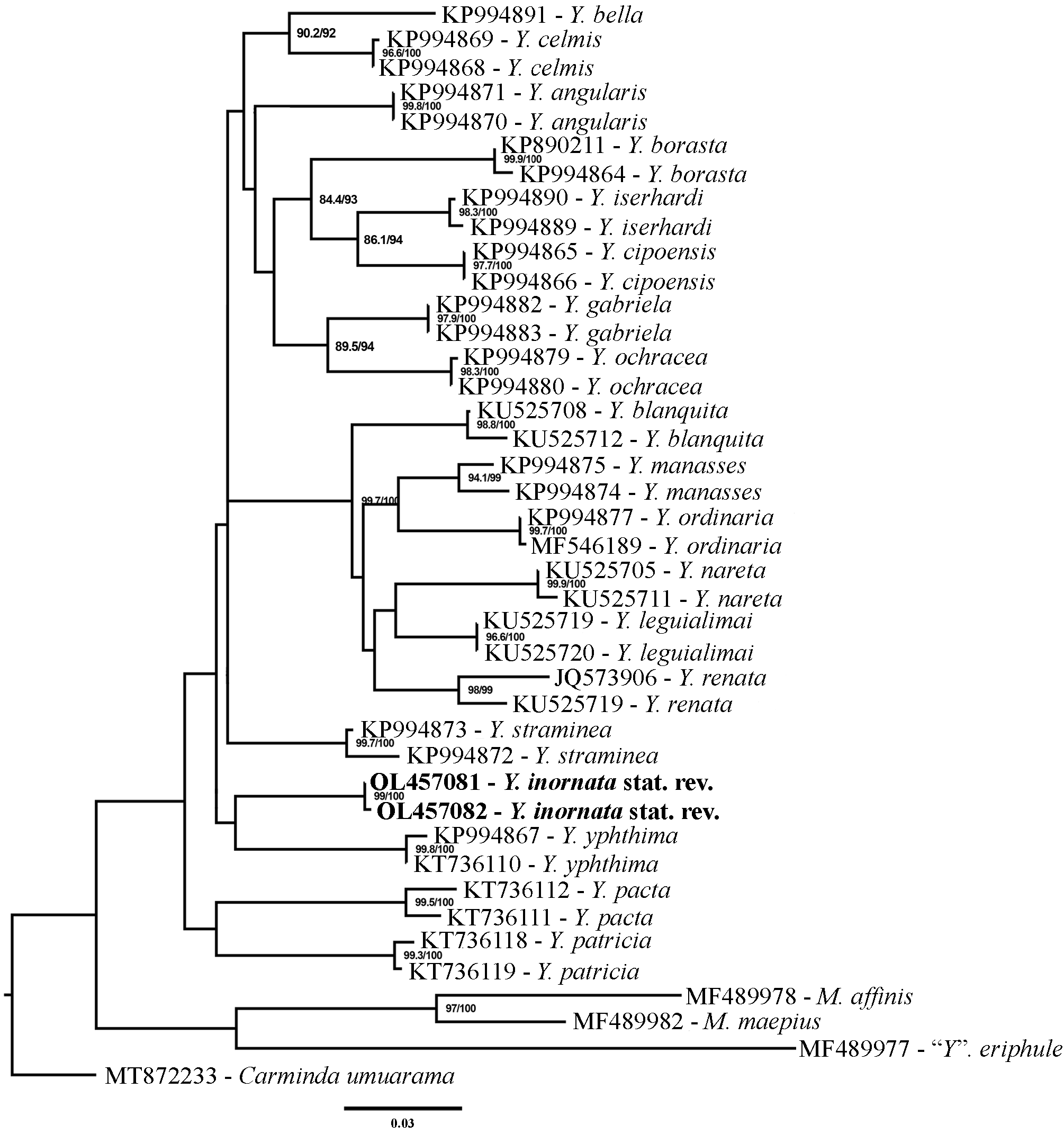
Diagnosis. Yphthimoides inornata stat. rev. differs from its congeners in the following combination of characters: Male on the dorsal forewing with a dark brown triangular androconial patch from CuA 1 to the inner margin ( Fig. 2A View FIGURE 2 ); male and female on dorsal hindwing with an eyespot in CuA 1 –CuA 2; on ventral forewing with an eyespot in M 1 –M 2; on ventral hindwing with eyespots in M 1 –M 2 and in CuA 1 –CuA 2, may have small ones in Rs–M 1 and in M 2 –M 3, varying in size ( Fig. 2 View FIGURE 2 ); ventral fore and hindwings without discal and postdiscal lines ( Figs 2B, 2D View FIGURE 2 ). In the molecular analysis Y. inornata stat. rev. is closely related to Y. yphthima but without support ( Fig. 8 View FIGURE 8 ); the genetic distance between them ranged from 6% to 7% ( Table 1 View TABLE 1 ).
......continued on the next page
Asterisk (*) indicates sequences provided by this study.
Immature stages. Egg ( Fig. 5A View FIGURE 5 ). Oviposition. 27–II–2019. Duration 7 days. Diameter 1.1 mm. Spherical, cream, with thin ridges forming a pattern of irregular pentagonal and hexagonal cells.
First instar ( Figs 5B, 5C View FIGURE 5 ). Duration 9 days. Head capsule width 0.7 mm; head hypognathous, dark brown with crackled aspect, covered by dark setae with creamy tips, and with two short horns on vertex, lateral adfrontal sutures and epicranial suture bear small circular-shaped sclerotizations; stemma 3 larger than the other stemmata; ventral margin of labrum moderately notched; head horn 0.08 mm.
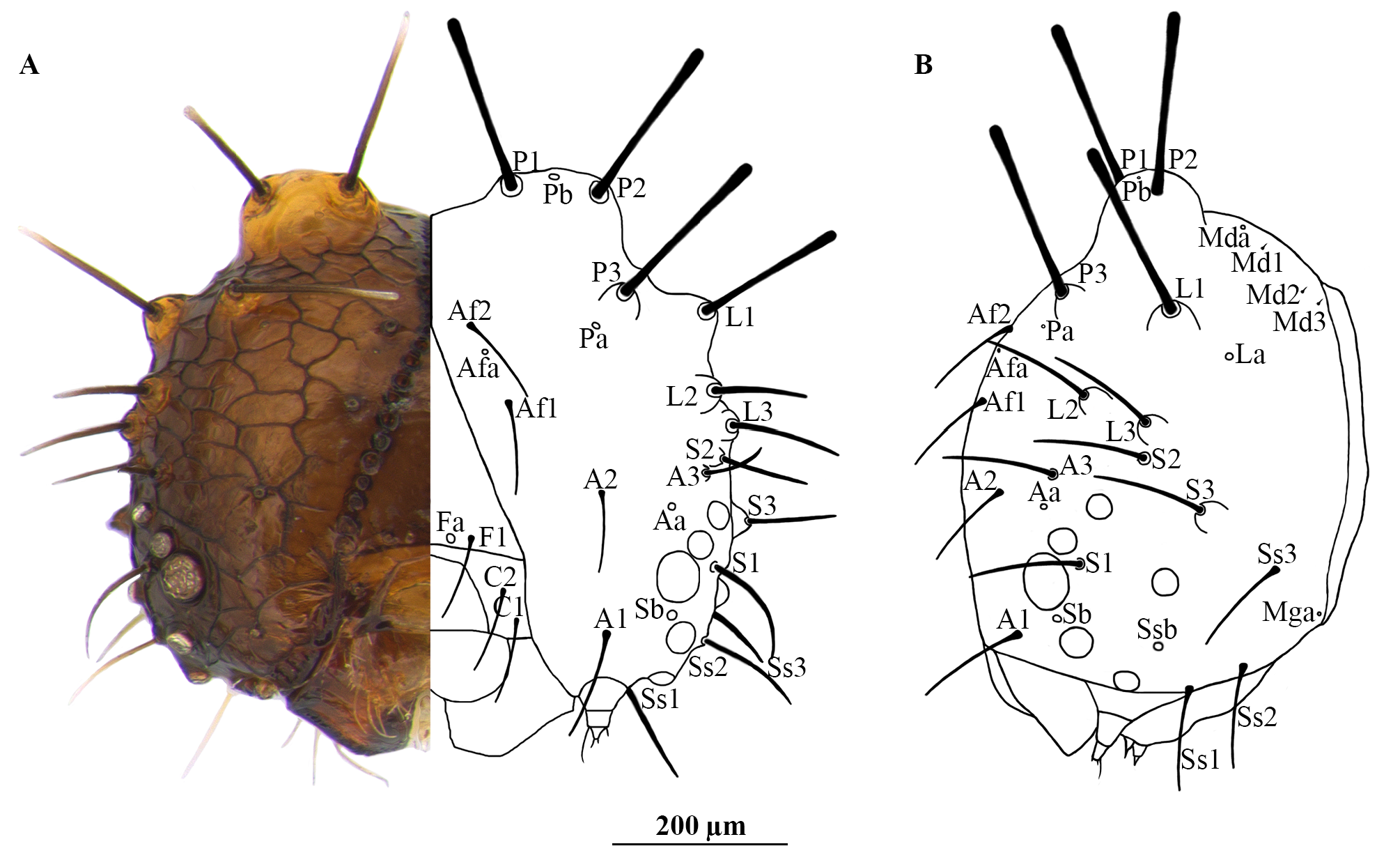
Head chaetotaxy ( Fig. 6 View FIGURE 6 ). L1 and P setae blunt at their tips, the other setae acute at their tips. One pair of frontal setae (F1), between them one pair of frontal pores (Fa); two pairs of adfrontal setae (Af1, Af2), between them one pair of adfrontal pores (Afa); on clypeus two pairs of setae (C1, C2); three anterior pairs of setae (A1, A2, A3) and one pair of pores (Aa); three lateral setae (L1, L2, L3) (L2 and L3 are proposed here), L3 near S2 and one pair of pores (La); three stemmatal setae (S1, S2, S3), and one pair of pores (Sb); three substemmatal setae (Ss1, Ss2 and Ss3) and one pair of pores (Ssb); one pair of microgenal setae (Mg1) at the ventrolateral occipital margin and one pair pores (Mga); three pairs of posterior setae (P1, P2, P3 Freitas et al. 2012, Cosmo et al. 2014), and two pairs of pores (Pa, Pb), Pb between P1 and P2; three pairs of microdorsal setae (Md1, Md2, Md3) and one pair of pores (Mda) above Md1.
Body length 4 mm; covered by cream setae, elongated, and clubbed at the tip; cream, with reddish brown longitudinal lines: the median-longitudinal line wider than subdorsal, supraspiracular and spiracular lines; a very short bifid ‘tail’ on the 10th abdominal segment.
Second instar ( Figs 5D, 5E View FIGURE 5 ). Duration 7 days. Head capsule width 1.03 mm; grayish brown with smaller setae than the first instar; head scoli 0.13 mm. Body length 6.6 mm; similar to the first instar but more grayish, with smaller setae, with the median-longitudinal line more evident and longer bifid ‘tail’.
Third instar ( Figs 5F, 5G View FIGURE 5 ). Duration 7 days. Head capsule width 1.48 mm; similar to the second instar but more brownish; with a cream subdorsal band from the head scoli tapering into the thorax and abdomen; head scoli 0.2 mm. Body length 11 mm similar to the second instar but more brownish, dorsally, between T1-T2 with a dark small spot; subdorsally, on T3 to A8, with dark small spot between each segment.
Fourth instar ( Figs 5H, 5I View FIGURE 5 ). Duration 8 days. Head capsule width 2 mm; head scoli 0.2 mm; similar to the third instar. Body length 18 mm, similar to the third instar.
Fifth instar ( Figs 5J, 5K View FIGURE 5 ). Duration 11 days. Head capsule width 3.6 mm; head scoli 0.3 mm; similar to the fourth instar, but the cream subdorsal band, with the same width from the head scoli to A10. Body length 38 mm, similar to the fourth instar.
Pupa ( Figs 6A–6C View FIGURE 6 ). Duration 13 days. Length 14 mm, integument smooth without ornamentation, ventrally lighter than dorsally; median dorsal line dark brown and more evident on the mesothorax, reddish brown from the metathorax to the cremaster. Head with elliptical eyes; triangular labrum with a dark brown spot; antenna and galea with the same length, ending on A4, and longer than mesothoracic leg. Thorax on the fore and mesothoracic legs with a dark brown spot each; alar theca with marked veins, ending in small black spots.Abdomen with several parallels longitudinal lines; A1 and A4 with two small dark spots each.
Distribution and etology. Yphthimoides inornata stat. rev., based on the examined material, including the holotype and the allotype, has the following geographical distribution: Brazil: Paraná (Guarapuava), Santa Catarina (Curitibanos, Joaçaba, São Bento do Sul), Rio Grande do Sul (Carazinho, Pelotas), and Argentina: Corrientes (Playadito), flying between February–May, and September–November, suggesting a multivoltine species. Adults were found flying close to the ground, in open grassland areas near forest fragments, during the hottest hours of the day.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |