
Mummucia ibirapemussu, Carvalho, Leonardo S., Candiani, David F., Bonaldo, Alexandre B., Suesdek, Lincoln & Silva, Paulo Roberto R., 2010
|
publication ID |
https://doi.org/ 10.5281/zenodo.199513 |
|
DOI |
https://doi.org/10.5281/zenodo.6212081 |
|
persistent identifier |
https://treatment.plazi.org/id/D90F6546-FFA2-7D4E-FF4D-FB30FDB8FEA0 |
|
treatment provided by |
Plazi |
|
scientific name |
Mummucia ibirapemussu |
| status |
sp. nov. |
Mummucia ibirapemussu View in CoL , new species
Figs. 2–31 View FIGURES 2 – 8 View FIGURES 9 – 12 View FIGURES 13 – 18 View FIGURES 19 – 24 View FIGURES 25 – 26 View FIGURE 27 View FIGURE 28 View FIGURE 29 View FIGURES 30 – 31 .
Type material. Holotype: Male from Serra das Confusões National Park (S09°03’85’’, W043°46’31’’), Guaribas and Caracol, Piauí, Brazil, VI–VII.2007, P.R.R. Silva et al. leg., MPEG 0 0 0 5. Paratypes: fifteen males and three females, same data as holotype, MPEG 0006-0015, IBSP and MZUEFS.
Etymology: From Tupi, an ancient Indian language, by agglutination of ibirapema = indigenous weapon used to kill a prey and uassu = large. This name makes reference to the large chelicerae of Solifugae .
Diagnosis: Mummucia ibirapemussu n. sp. resembles M. mauryi by the pleurite coloration but differ to that species by its movable finger teeth, graded in size from distal to proximal II, III and I ( Figs. 9–10 View FIGURES 9 – 12 ; 13–14). Besides, Mummucia ibirapemussu n. sp. resembles Metacleobis fulvipes concerning the overall cheliceral dentition. In Mummucia ibirapemussu n. sp, the fourth distal tooth of fixed finger is smaller than the second (males and females) whereas in Metacleobis fulvipes the size of those teeth are similar. Total body length is smaller in Mummucia ibirapemussu n. sp. Pleurite coloration is different between these two species.
Description. Male holotype (MPEG 0005).
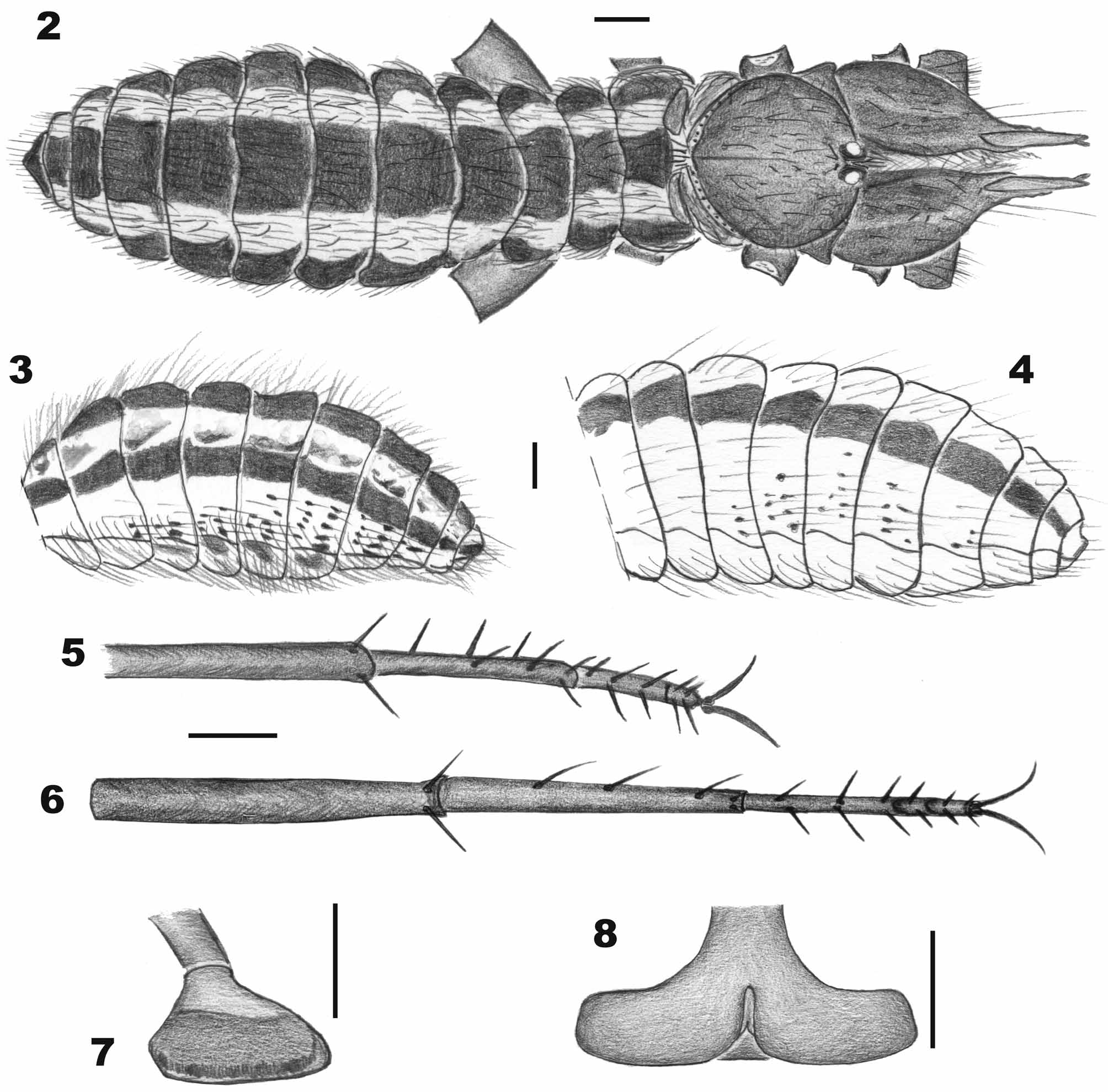
Coloration in 70% ethanol. Prosoma: propeltidium whitish yellow, with central portion pale yellow larger than ocular tubercle in the median region, whitish yellow in the border of the lateral lobes. Ocular tubercle with a narrow longitudinal whitish yellow stripe between the eyes and a black ring surrounding each eye. Peltidium whitish yellow and parapeltidium white. Mesopeltidium and metapeltidium with a brown central stripe and lateral white stripes, each stripe a third of the tergite width ( Fig. 27 View FIGURE 27 ).
Chelicerae pale yellow, with three longitudinal, distally fused, white stripes on the ectal face.
Pedipalpi and legs pale yellow, with slightly darker ends. Malleollus of the trochanter II grey with large brown spots on the enlarged portion ( Figs. 7 View FIGURES 2 – 8 , 17 View FIGURES 13 – 18 ) and the remaining malleollus whitish.
Opisthosoma: lateral borders of the tergites white, with a wide dark brown stripe on the central half, which is darker near the posterior border of the tergites. White area of the tergites covered by brown bifid setae with brown sockets and scattered brown marks. Pleurites white on the ventral portion, dorsal portion brown. Pale brown translucent bifid bristles in the white portion have sockets shaped into brown spots. Sternites white, lateral borders brownish. Anal segment almost totally brown. First to fourth postspiracular sternites with brown spots which include the sockets of some bifid bristles. All covering bristles and bifid bristles are translucent yellowish.
Morphology and chaetotaxy. Prosoma: propeltidium wider than long (Table 1) with some scattered bifid setae, and separated from lateral lobes by dorsal grooves. Ocular tubercle prominent with one transversal row of anteriorly oriented bifid setae and two longitudinal rows between the eyes. Distance between two eyes about 1.5 times eye diameter. Peltidium narrow, with a transverse row of different sized bifid setae. Parapeltidium smooth. Mesopeltidium trapezoidal-shaped, 2.5 times wider than long, with one row of bifid setae on posterior margin. Metapeltidium 1.6 times wider than long.
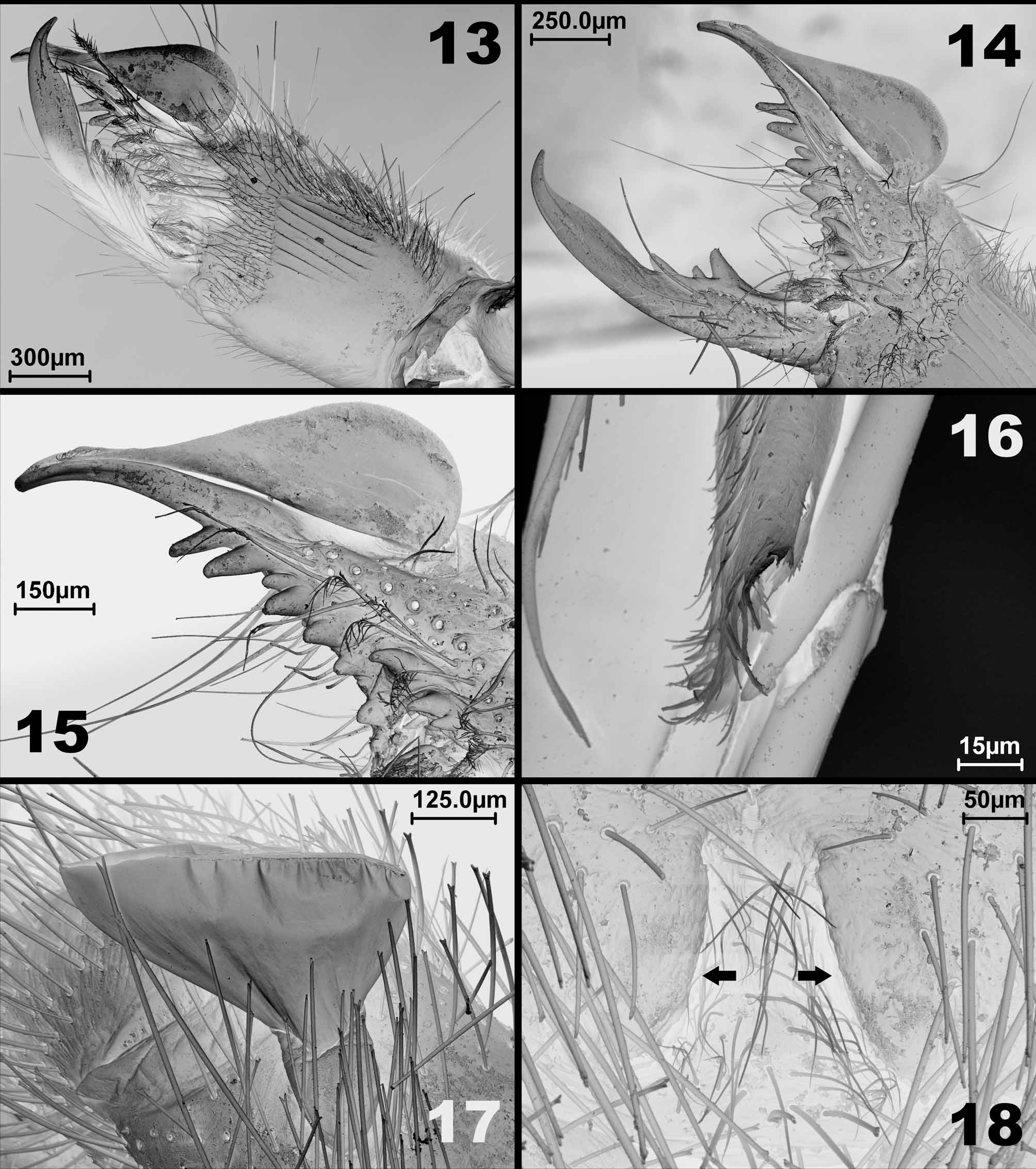
Chelicerae ( Figures 9–10 View FIGURES 9 – 12 ; 13–15): stridulatory apparatus on mesal face with seven parallel grooves ( Fig. 13–14 View FIGURES 13 – 18 ). Ectal face with several short bristles and setae, both bifid and acuminate. Movable finger with one anterior, one intermediate, and one principal tooth, graded in size from distal to proximal II, III and I ( Figs. 9– 10 View FIGURES 9 – 12 , 14 View FIGURES 13 – 18 ). Fixed finger with two anterior and two principal teeth graded in size from distal to proximal II, I, IV, III; five ectal fondal teeth, graded in size: I, III, V, II, IV and three mesal fondal teeth, graded in size from distal to proximal I, II, III, the first distal separated from the others by a diastema ( Figs. 9–10 View FIGURES 9 – 12 , 14–15 View FIGURES 13 – 18 ). Fixed finger bears, centrally on its dorsal face, one very long seta (about the length of femur III) with a prominent socket ( Fig. 14 View FIGURES 13 – 18 ).
Flagellum ( Figs. 9–10 View FIGURES 9 – 12 , 13–16 View FIGURES 13 – 18 ): a thin translucent drop-shaped vesicle, laterally flattened as in other mummuciids; with a long longitudinal ectal opening. The attachment of the flagellum is a sclerotized ring placed posteriorly on its ectal face; and its distal portion is irregularly brush-like ( Fig. 16 View FIGURES 13 – 18 ).
Pedipalp: tarsi immovable, without spines, densely covered by different sized bifid bristles and several short acuminate bristles, with some very long setae in tibiae and femur (about 1.1 times pedipalpal tibial length).
Legs: covered by several different sized bifid bristles and some bifid setae (see Fig. 26 View FIGURES 25 – 26 ). Some very long setae on the dorsal surface (about 2.6 times the basitarsus IV length). Leg I: thin, without claws and spines. Legs II and III: tibiae with a distal pair of ventral spines; basitarsus with three retrolateral spines and 1.1.2 ventral spines; telotarsi two-segmented with 1.2.2/2.2 ventral spines ( Fig. 5 View FIGURES 2 – 8 ). Leg IV: tibia with a distal pair of ventral spines; basitarsus with 1.1.1.2 ventral spines; telotarsi three-segmented, with 2.2.2/2/2.2 ventral spines ( Fig. 6 View FIGURES 2 – 8 ). The three segments of the telotarsi IV with some ventral spines can be seen in Figure 26 View FIGURES 25 – 26 . Malleoli as in Figures 7 View FIGURES 2 – 8 and 17 View FIGURES 13 – 18 .
Opisthosoma: tergites wider than long ( Figs. 2 View FIGURES 2 – 8 , 27 View FIGURE 27 ), with rounded borders, covered by bifid setae and bifid bristles. Genital operculum with central longitudinal opening. Sternites wider than long, densely covered by bifid bristles. Posterior border of 2nd post-spiracular sternite with a row of about 50 ctenidia, more rigid and slightly longer than the bifid bristles in sternites ( Fig. 24 View FIGURES 19 – 24 ). Two post-spiracular sternites with several long and acuminate bristles, 1.5 times longer than other sternite bristles. Pleurites as in Figure 3 View FIGURES 2 – 8 .
Measurements: see Table 1.
Female paratype (MPEG 0010).
As in male, except for the following features.
Morphology and chaetotaxy. Prosoma: Eyes separated by 1.25 times their diameter. Mesopeltidium twice wider than long. Metapeltidium 1.7 times wider than long.
Chelicerae ( Figs. 11–12 View FIGURES 9 – 12 , 19–22 View FIGURES 19 – 24 ): Movable finger with one anterior, one intermediate, and one principal tooth, graded in size from distal to proximal II, III, I ( Figs. 11–12 View FIGURES 9 – 12 , 20, 22 View FIGURES 19 – 24 ). Fixed finger with two anterior and two principal teeth graded in size from distal to proximal II, I, IV, III; five ectal fondal teeth, graded in size: I, III, V, II, IV ( Figs. 11–12 View FIGURES 9 – 12 , 20–21 View FIGURES 19 – 24 ). Three (as in males; Fig. 12 View FIGURES 9 – 12 ) or two mesal fondal teeth separated by a diastema.
Opisthosoma: sternites densely and uniformly covered by bifid bristles, without conspicuous sockets ( Fig. 24 View FIGURES 19 – 24 ). Genital operculum ( Fig. 23 View FIGURES 19 – 24 ) prominent, round-bordered, with central longitudinal opening. Pleurites as in Figure 4 View FIGURES 2 – 8 .
Measurements: see Table 1.
Intraspecific variation. The white area of the tergites of most of the male paratypes lack the brown scattered marks present on the holotype, probably owing to degradations of the tissue caused by poor preservation of the specimen. The female used for SEM pictures had only four ectal fondal teeth, while the remaining female paratypes have five ectal fondal teeth.
TABLE 1. Morphometric characters of Mummucia ibirapemussu n. sp.. Measurements are in millimeters
(except propeltidium length/width ratio).
Remarks. The new species herein described, Mummucia ibirapemussu n. sp., is assigned to the type genus Mummucia , because at the present time it is impossible to reliably distinguish the genera of Mummuciidae , as pointed out by Maury (1998). The same decision was taken by Xavier and Rocha (2001), Martins et al. (2004) and Rocha and Carvalho (2006). The family Mummuciidae still requires an entire taxonomic and phylogenetic revision to better understand the generic limits and define the most useful characters for species definition. However, one character appears to be constant in the family: the longitudinal ectal opening of the flagellum, which was already reported only for Mummucia coaraciandu , M. mauryi , M. taiete Rocha and Carvalho, 2006 ( Rocha and Carvalho, 2006: 165, fig. 5) and Metacleobis fulvipes ( Xavier and Rocha, 2001; Martins et al. 2004; Rocha and Cancello 2002b; Rocha and Carvalho, 2006).
Mummucia ibirapemussu View in CoL n. sp. slightly resembles M. mauryi View in CoL in the color pattern of pleurites, a character set that was proposed to be species-specific in Mummuciidae ( Rocha and Carvalho, 2006) View in CoL . This feature appears to be convergent in these two species, as they differ in the cheliceral dentition, a character used by Maury (1984) and Rocha (2002) to support species recognition.
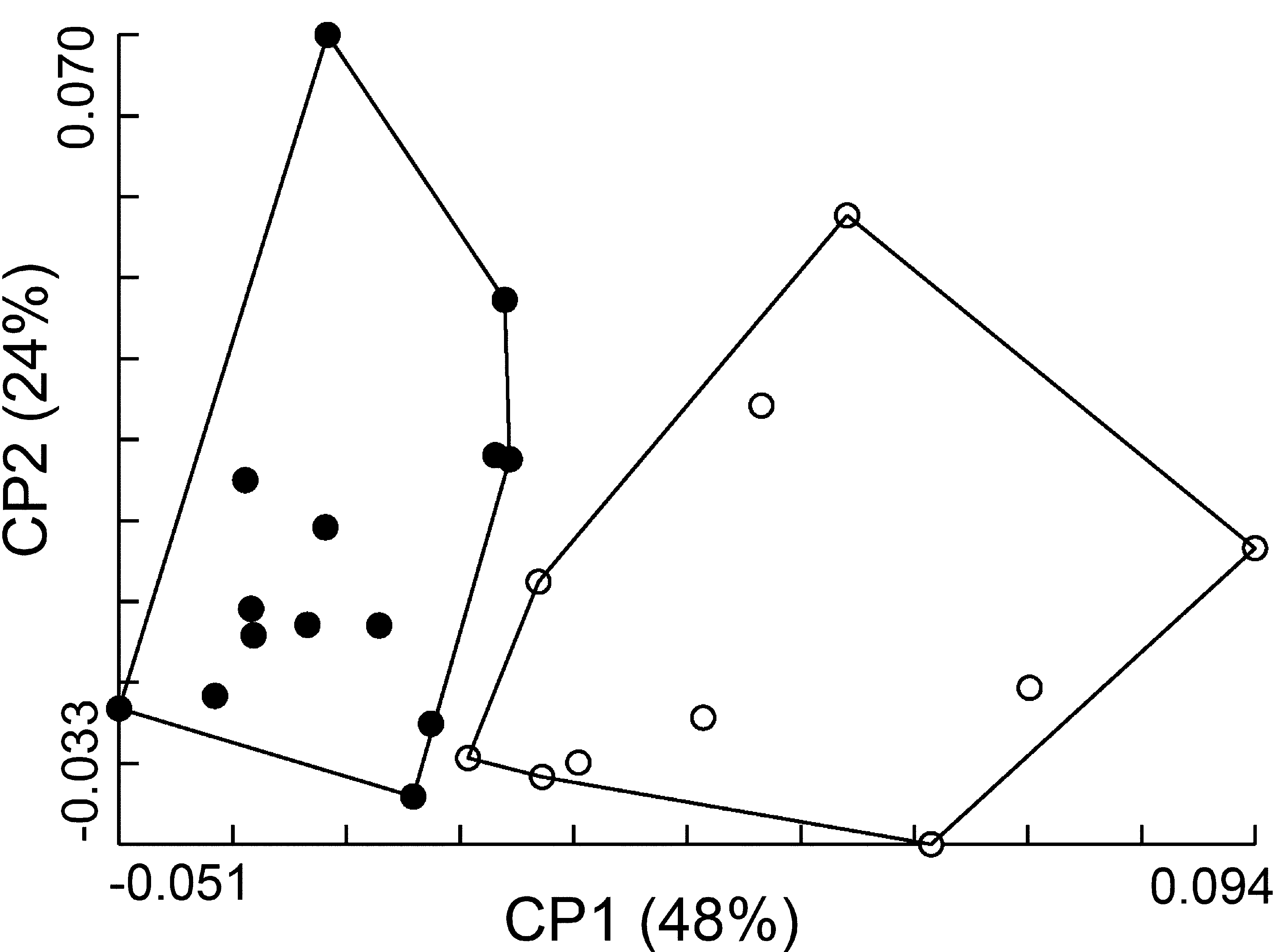
Besides, the geometry of propeltidium showed that males of M. ibirapemussu View in CoL n. sp. can be biometrically distinguishable from Metacleobis fulvipes View in CoL . Shape analyses showed the individuals arranged in distinct plot groups at the graphical morphospace defined by principal components 1 and 2 ( Fig. 28 View FIGURE 28 ). Each group corresponded to one species, with no overlap between them. The reclassification accuracy of females based on the Mahalanobis distances was 100% for M. ibirapemussu View in CoL n. sp. and 92% for Metacleobis fulvipes View in CoL .
To help in distinguishing M. ibirapemussu View in CoL n. sp from Metacleobis fulvipes View in CoL , the morphometric characters of propeltidium shape were examined from a multivariate point of view. Shape of propeltitium is taxonomically informative and for that reason it has been frequently addressed in species descriptions, represented by its length/width ratio ( Muma, 1951; Maury, 1982, 1987; Rocha and Carvalho, 2006).
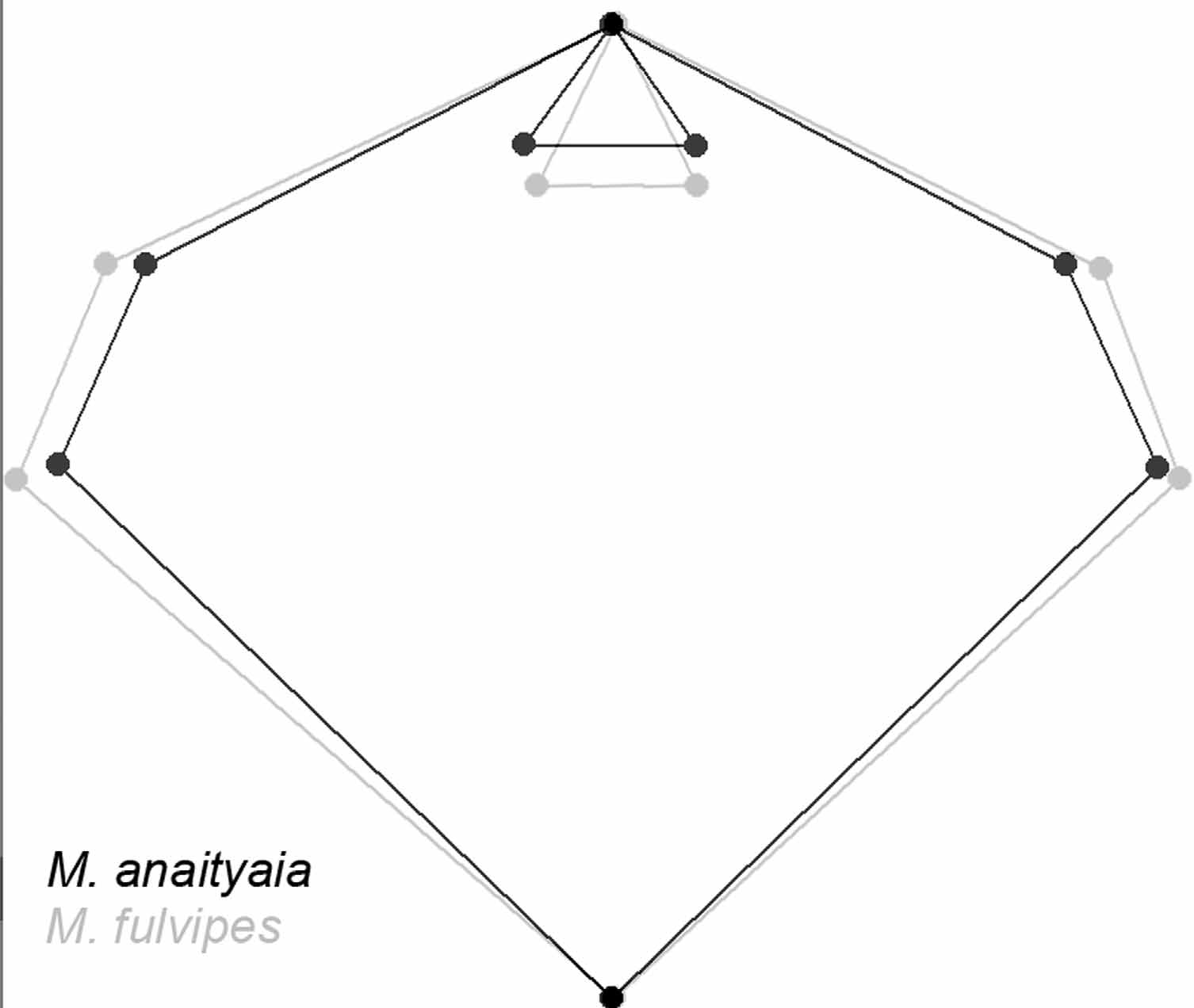
Superimposition of consensus configurations showed that LMs 2–8 positioning are distinct between both species ( Fig. 29 View FIGURE 29 ). Mean values of ratio propeltidium length/width were: M. ibirapemussu View in CoL n. sp. =0.90; Metacleobis fulvipes View in CoL =0.81. Another morphometric character that appears to be diagnostic for these species is the ratio obtained through the division of propeltidium length by the arithmetic mean of the distances LM1- LM7 and LM1-LM8. Such ratio for each species was: M. ibirapemussu View in CoL n. sp. =6.31; Metacleobis fulvipes View in CoL =5.34.
Geometric morphometrics was accurate to diagnose two species Solifugae View in CoL , similarly as observed in Insecta, a group in which several cryptic species has been characterized by such method ( Dujardin, 2008). Arguably, morphometrics could be helpful for distinguishing other species or genus of Solifugae View in CoL when larger taxonomic samples are available.
Habitat preference. We collected fourteen specimens (eleven males and three females), all in July 2007, when the enclave (humid area formed between hills) was not sampled. A total of twelve individuals were collected in arboreal Caatinga ( Fig. 30 View FIGURES 30 – 31 ) and seven individuals in shrubland Caatinga ( Fig. 31 View FIGURES 30 – 31 ). The low number of individuals sampled and the punctual sampling events prevent us to infer about habitat selection for Mummucia ibirapemussu View in CoL n.sp.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Mummucia ibirapemussu
| Carvalho, Leonardo S., Candiani, David F., Bonaldo, Alexandre B., Suesdek, Lincoln & Silva, Paulo Roberto R. 2010 |
Mummuciidae (
| Rocha and Carvalho 2006 |