
Hydrocyphon kopetdaghensis, Ruta, Rafal, 2007
|
publication ID |
https://doi.org/ 10.5281/zenodo.177443 |
|
DOI |
https://doi.org/10.5281/zenodo.6243983 |
|
persistent identifier |
https://treatment.plazi.org/id/D97187B8-A322-FF91-1AAB-10476AA3FC22 |
|
treatment provided by |
Plazi |
|
scientific name |
Hydrocyphon kopetdaghensis |
| status |
sp. nov. |
Hydrocyphon kopetdaghensis sp. nov.
( Figs. 1–7)
Holotype. USSR, Turkmenia, Kopet-Dagh / Mts., 1000 m, Kurkulab, 6 km W / Germob, 57º50’E, 38º04’N / 0 3.10.1991, No. L34 / leg.: A. Podlussány, L. Ronkay & Z. Varga [white label, printed]. HYDROCYPHON kopetdaghensis sp. nov. / des. Rafał Ruta, 2007 / HOLOTYPUS [red label, printed]. Deposited in Hungarian Museum of Natural History.
Diagnosis. Easily distinguishable on the basis of genital characters: the shape of parameres combined with structural anatomy of the penis are unique for this species of Hydrocyphon .
Description. Male. Body oval, slightly depressed, shiny, clothed with bright, yellowish-white hairs. Elytra brownish, pronotum yellow with darkened disc, head dark brown. Venter brown. Legs yellow with slightly darkened tarsi. Maxillary palpi yellowish-brown. Antennomeres 1-4 yellow, 5 brown, 6-11 dark brown. Length (measured from the anterior edge of pronotum to the apex of elytra) 2.0 mm, greatest width of pronotum 0.9 mm, greatest width of elytra 1.4 mm, greatest depth of body 0.7 mm, body 1.46 times as long as broad.
Greatest width of the head 0.65 mm. Head 1.86 times as broad as long, 1.6 times wider than width of interocular space, with distinct punctation, and strongly protuberant eyes. Antenna filiform; length ratio of antennomeres: 2.9: 2.3: 1.0: 2.5: 2.1: 2.1: 2.1: 2.1: 2.1: 2.1: 2.1: 3.9; length/width ratio of antennomeres: 1.6, 1.3, 1.1, 1.8, 1.5, 1.5, 1.5, 1.5, 1.5, 1.4, 2.9. Anterior clypeal margin straight, with rounded anterior angles. Labrum with rounded margin. Mandibles reduced, with obtuse apices.
Pronotum small, 2.57 times as broad as long, widest at its base. Disc of pronotum with subtle punctation. Posterior pronotal margin almost straight, slightly sinuate. Lateral margins straight, slightly converging anteriorly. Anterior angles not produced.
Scutellum triangular, as long as broad, with indistinct punctation. Base of elytra evenly wider than base of pronotum, sides rounded. Elytra without longitudinal ridges, with marked humeri, 1.32 times as long as broad and 5.58 times as long as pronotum. Subtle adsutural strioles are present in proximal half of elytra. Punctation relatively dense, punctures separated by a single diameter. Punctures larger than those on pronotum. Epipleura yellow, gradually narrowing, virtually obsolete in subapical region of elytra. Hind wings fully developed.
Prosternal process elongated, very narrow, rod-like. Mesocoxae separated by a long subrectangular process (ca. 3 times as long as wide), metaventral discrimen present in posterior 2/3 of metaventrite. Metaventral process reduced, subtriangular, metacoxae touching one another. Five abdominal sternites visible. Ratio of ventrites lengths: 1: 2: 2: 2: 2.5. Right metafemur of the holotype is underdeveloped – shorter than that of the left.
Genitalia. Penis symmetrical, with long, regularly curved parameroids and wide prosthemes; both parameroids and prosthemes somewhat up-curved; length (L) = 0.64 mm., width (W) = 0.24 mm. Tegmen with parameres wide, produced apically into sharp, excurved hooks; L = 0.52 mm, W = 0.34 mm. Tergite VIII with a row of minute setae on apical margin; L = 0.41 mm, W = 0.35 mm. Tergite IX membranous in central part of apical margin; L = 0.36 mm, W = 0.32 mm. Sternite VIII v-shaped, L = 0.25 mm, W = 0.28 mm. Sternite IX consisting of two hemisternites, each of which consists of a long baculus, with widened apex bearing several setae; L = 0.46 mm, W = 0.24 mm (whole sternite).
Name derivation. Named after terra typica – Kopet Dagh Mountains.
Distribution. Known only from its locus typicus in Kopet Dagh Mountains (Turkmenia).
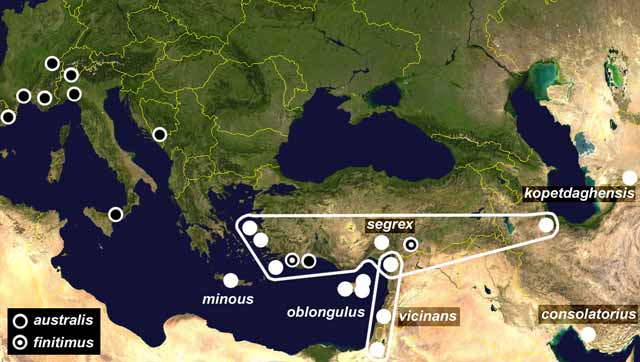
Discussion. On the basis of structural anatomy of the penis (both parameroides and prosthemes symmetrical) and tegmen (parameres wide), the species is classified within the Hydrocyphon australis -group, sensu Nyholm (1967) ( Klausnitzer 2002, Yoshitomi & Klausnitzer 2003), known from the eastern part of Western Palaearctic Region, with three species occurring in the Mediterranean Subregion, and four in the Middle East. Four of the five species of Hydrocyphon known to occur in the Middle East, belong to the australis -group. Their distribution is shown in Fig. 8 View FIGURE 8 (based on data from: Nyholm 1967, 1972, 1977; Klausnitzer 1970, 1981, 1989, 1990, 1990b, 1990c). An oriental Hydrocyphon bhutanensis Klausnitzer was included in the australis -group both in the original description ( Klausnitzer 1976) and in a recent check-list ( Yoshitomi & Satô 2005), although its placement in deflexicollis -group does not seem to be well supported. The main difference between these groups is the anatomy of penis—in the deflexicollis -group, the parameroids are symmetrical, in contrast to asymmetrical prosthemes; the structural anatomy of the tegmina is similar in both groups. Klausnitzer (1976) mentioned that H. bhutanensis has slightly asymmetrical prosthemes, a feature unusual in australis - group, nevertheless according to Klausnitzer, it may be regarded as a member of this group, with marked tendency to develop subtle asymmetry. In the author’s opinion, the species in question may also be regarded as a member of deflexicollis -group with prosthemes not as asymmetrical as usual in this group, and therefore artificially resembling species from the australis -group. Actually, a few oriental species closely resembling H. bhutanensis were recently included in the deflexicollis -group (e.g. Hydrocyphon lusonensis Yoshitomi & Satô, 2005 ; Hydrocyphon subcelatus Yoshitomi & Satô, 2005 ). The other hint to exclude H. bhutanensis from the australis -group is its geographical distribution. The australis -group seems to have its distributional centre in the Middle East and Eastern Mediterranean. The existence of a related species in the Oriental Region seem to be doubtful. It is highly plausible, that some of species-groups proposed in the most recent studies (especially the largest groups: pallidicollis -group and deflexicollis -group) are artificial assemblages of unrelated species.
Acknowledgements. I am greatly indebted to Dr. Ottó MERKL for allowing me to study the Scirtidae holdings of the Hungarian Museum of Natural History (Budapest, Hungary) and to Mr. Christopher Majka (Nova Scotia Museum of Natural History, Halifax, Canada) for linguistic assistance. My studies on Scirtidae were supported by Synthesys-grants (FR-TAF-890, BE-TAF-1969, AT-TAF-2276).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |