
Labulla, SIMON, 1884
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2005.00147.x |
|
persistent identifier |
https://treatment.plazi.org/id/D97887B6-FFF5-D97F-F7A5-CED49048F9E5 |
|
treatment provided by |
Diego |
|
scientific name |
Labulla |
| status |
|
LABULLA SIMON, 1884 View in CoL View at ENA
Labulla Simon (1884: 261) View in CoL . Type species: Linyphia thoracica Wider, 1834: 254 , pl. 17, f. 180. Although Simon described the genus in 1884, the type species was designated in Simon (1894: 707).
Diagnosis
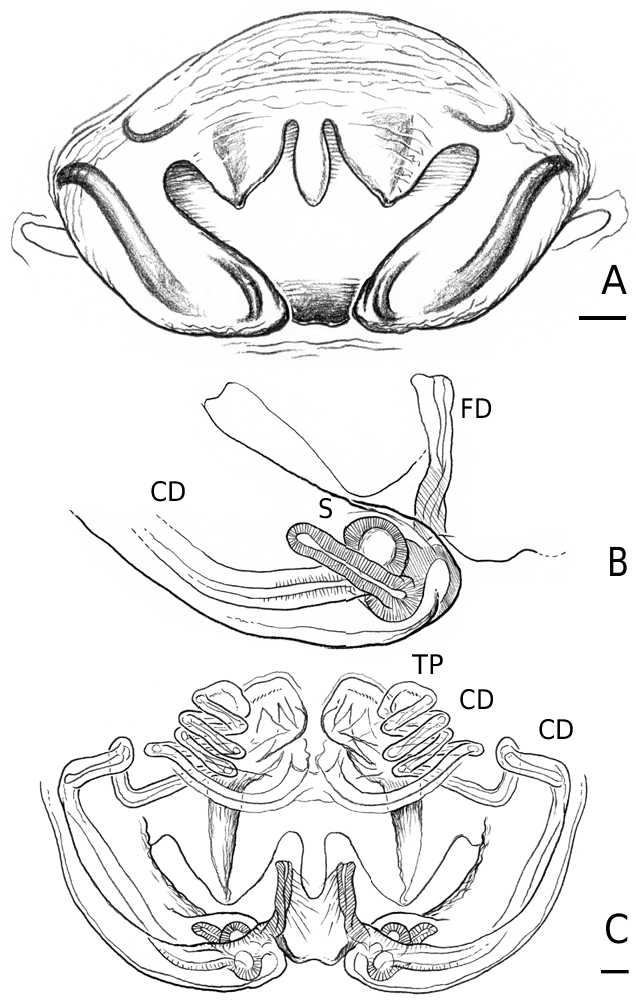
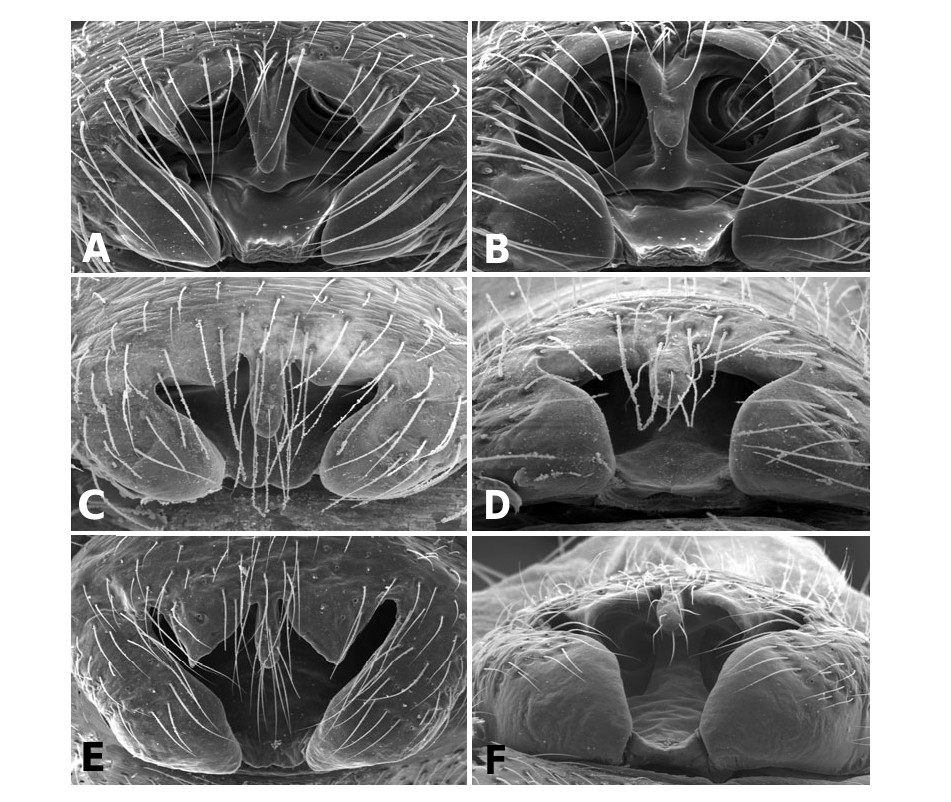
Male Labulla differ from other linyphiids by the following combination of characters: palpal tibia with dorsal process; paracymbium with a caudad apophysis in the medial branch, presence of an ectal tegular flap (membranous and running along longitudinal axis), presence of a bifid suprategular apophysis adjacent to the column; absence of embolic membrane and sheathlike lamella characteristica with a longitudinal fold housing distal region of the embolus. Females differ from other female linyphiids by the following combination of characters: presence in the epigynum of anterior flaps on both sides of a ventral plate, scape (without socket) and conspicuous posterior lateral lobes and U-shaped spermathecae with anteriorly orientated fertilization ducts. Males and females of all species have a patch of white spots ventrally on the abdomen, just in front of the spinnerets. This can be difficult or impossible to see in old preserved material.
Description
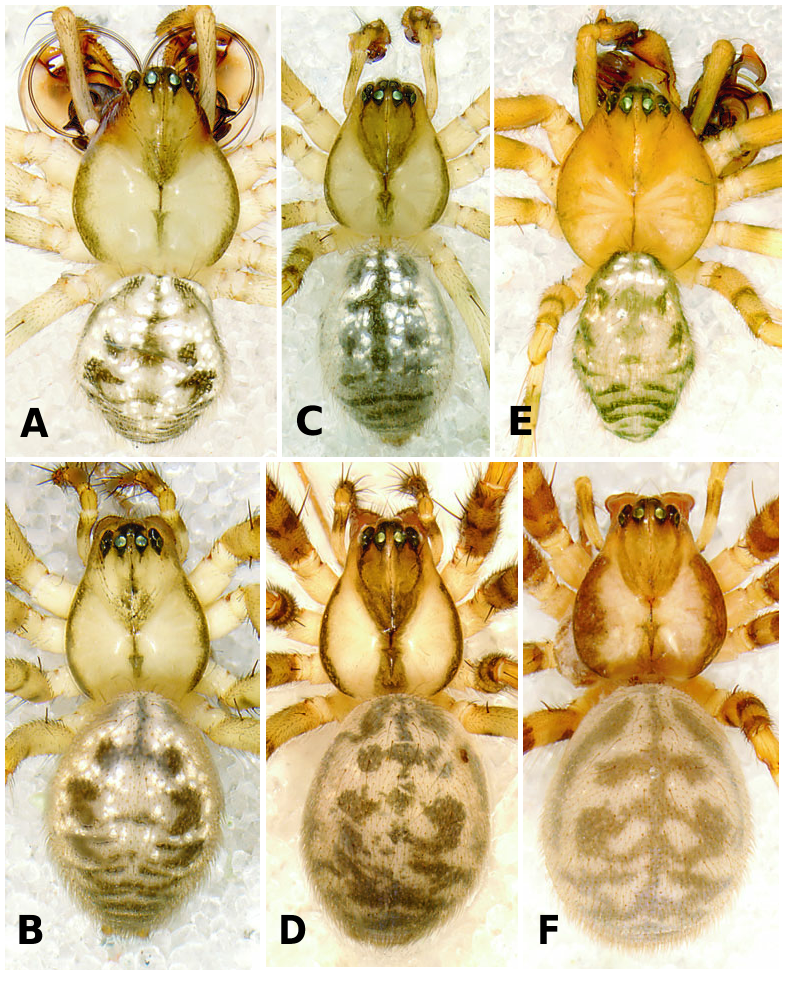
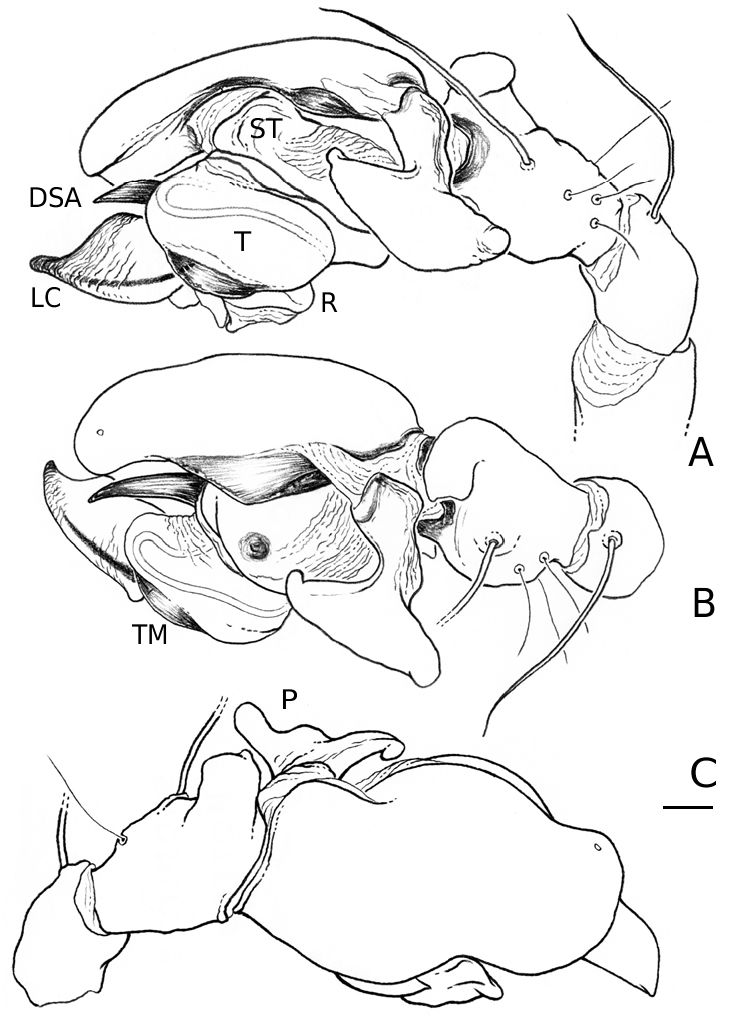
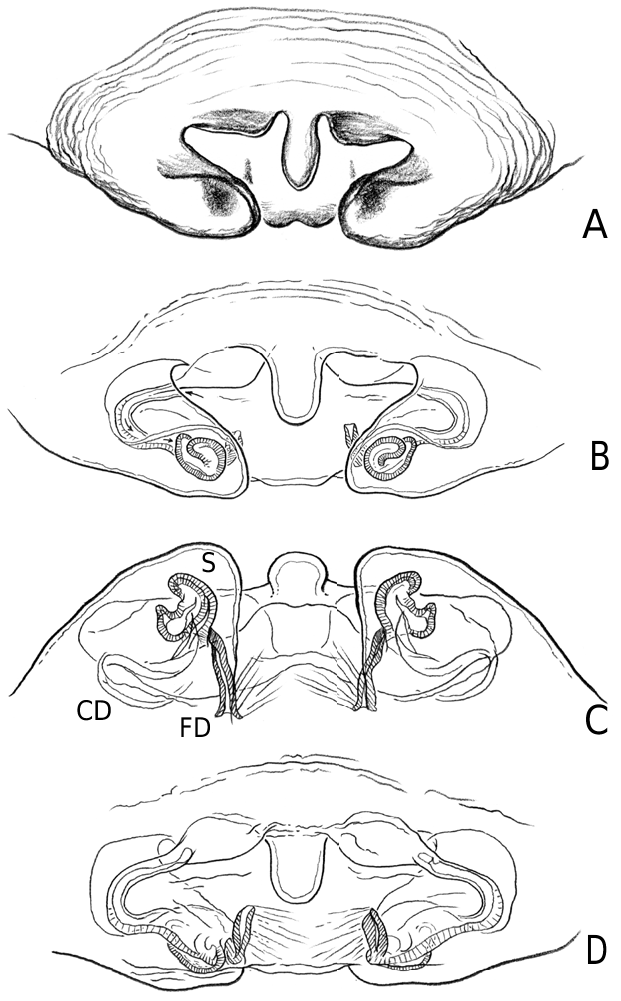
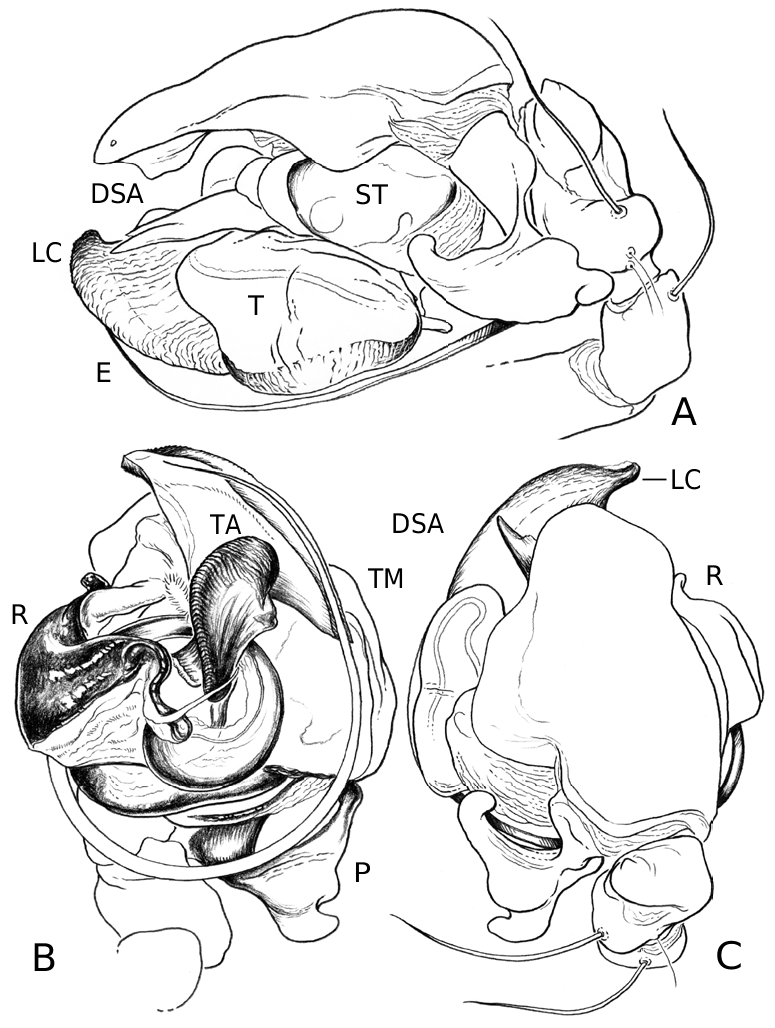
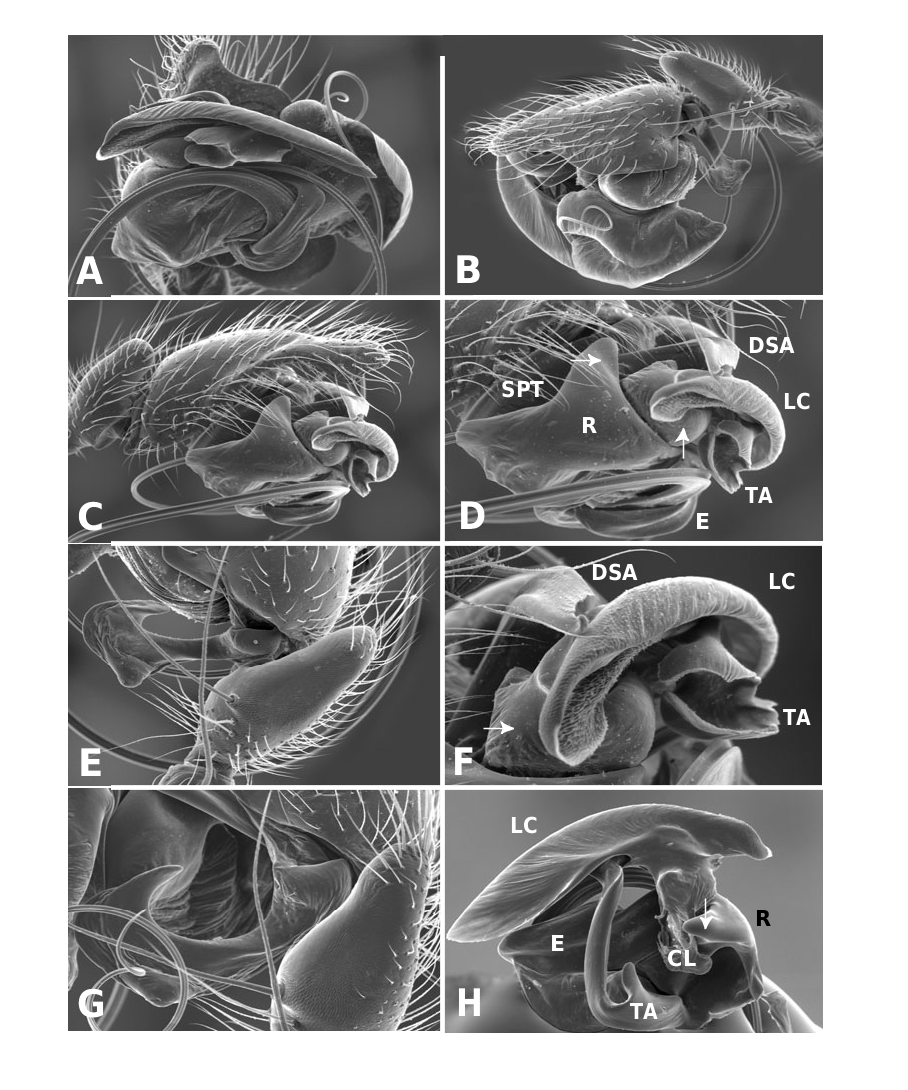
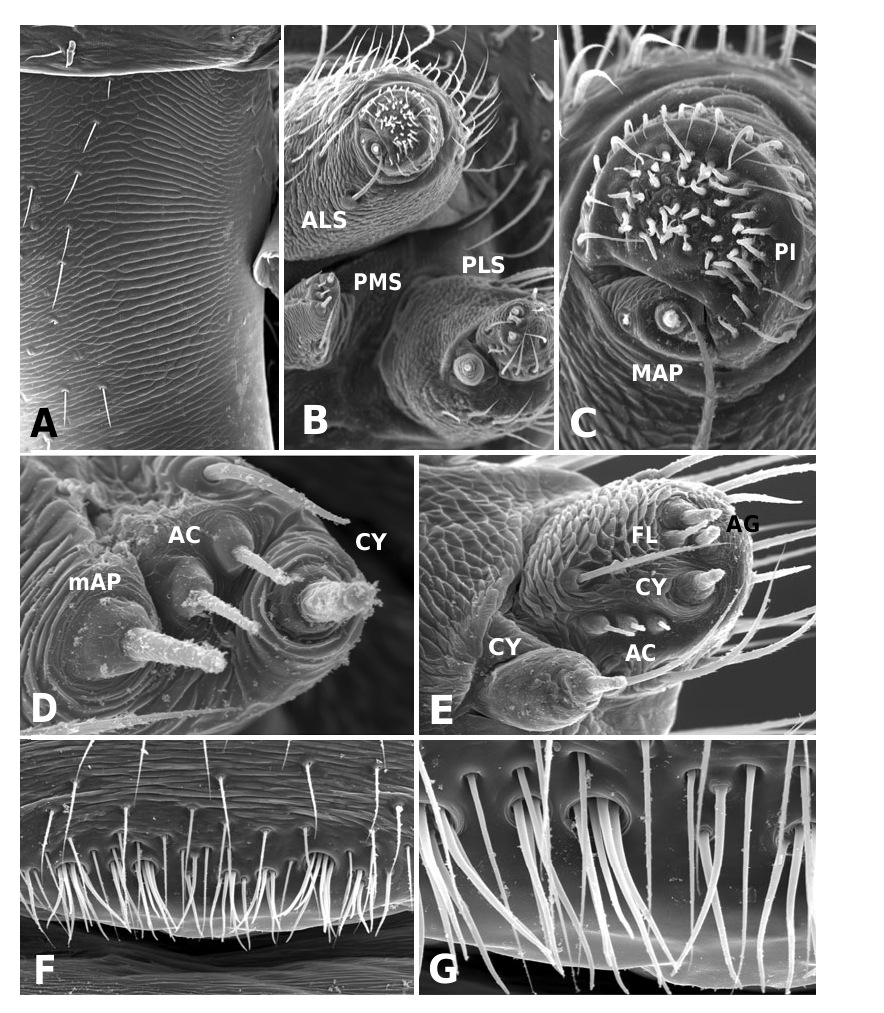
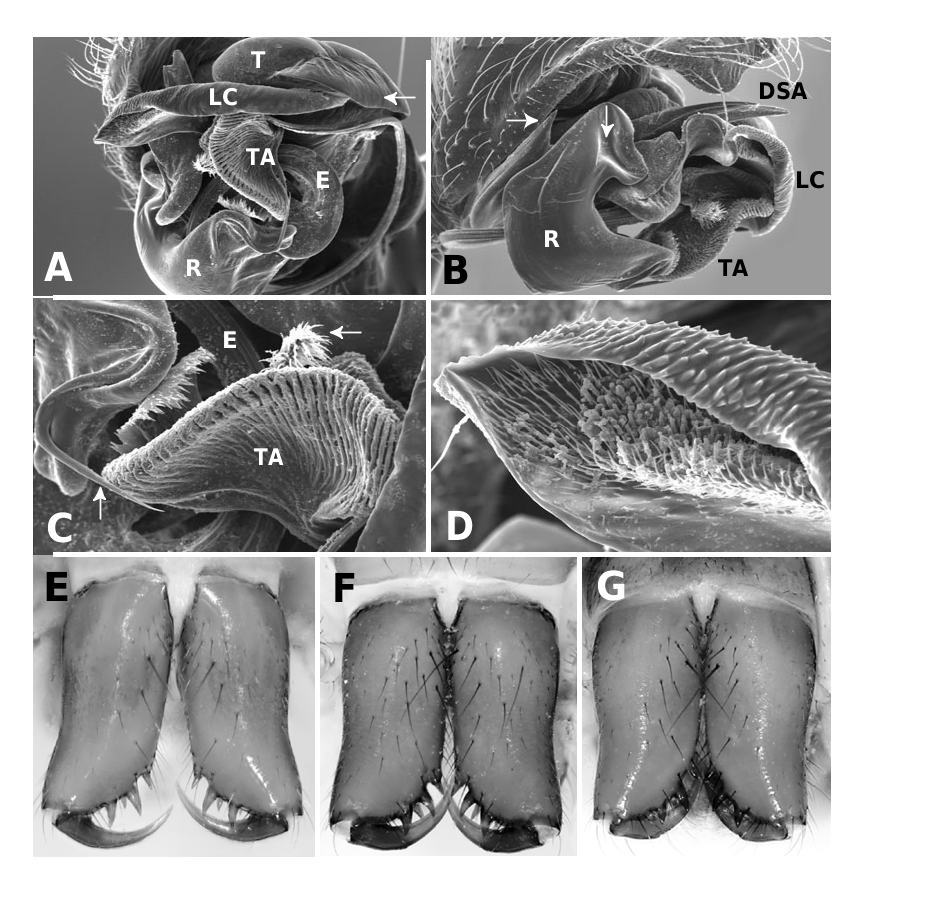
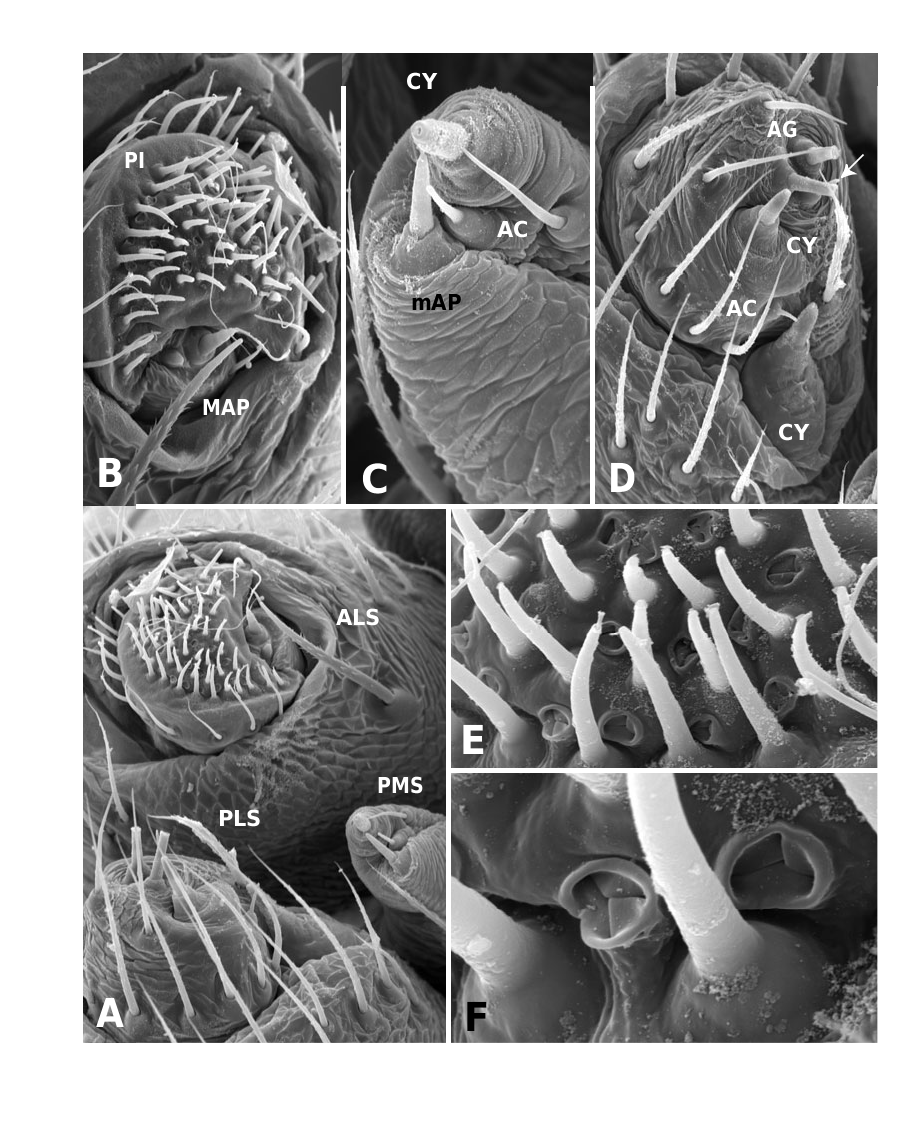
Medium to large linyphiid spiders, total length 4.00– 5.20 in males and 3.36–6.32 in females. Carapace longer than wide, piriform, 1.90–2.40 in males and 1.60–2.41 in females. Cephalothorax and chelicerae pale yellowish brown, carapace with greyish brown borders, with darker yellowish brown cephalic area (except male of L. machadoi sp. nov., Fig. 1 View Figure 1 ). Black rings around all eyes ( Fig. 1 View Figure 1 ). Thoracic furrow deep, running longitudinally from the posterior end of the cephalon to the posterior edge of the carapace. Sternum dark greyish brown. Legs yellowish brown, with dark annulations. Abdomen light grey, with dark grey pattern and white spots ( Fig. 1A View Figure 1 ). Ventrum with white dots, in variable pattern, between the book lungs and spinnerets (this can be difficult or impossible to see in old preserved material). Chelicerae with stridulatory striae. Clypeus height 5.75–2.23 times one AME diameter in males and 1.91–2.50 times one AME diameter in females. Chelicerae with three large widely spaced prolateral teeth and three to four small closely spaced retrolateral, proximal teeth ( Fig. 22E–G View Figure 22 ). Femur I 1.64–1.96 (males) and 1.27–1.76 (females) times the length of cephalothorax. Legs covered with numerous spines. Femur I–IV with one dorsal spine; femur I with additional prolateral spine. Tibia I–IV with two dorsal spines; all tibiae with additional ventral, pro- and retrolateral spines. Metatarsi I–IV with two dorsal spines; all metatarsi with additional pro- and retrolateral spines, but no ventral spines. Metatarsus I trichobothrium 0.2–0.3 (sometimes absent in male L. thoracica ). Metatarsus IV trichobothrium absent. Female palp with tarsal claw. Leg autospasy at the patella–tibia junction. Colulus large and fleshy, covered with short setae and two long setae (longer than the colulus itself) on each side. Spinnerets typical of a linyphiine ( Hormiga, 1994b, 2002) ( Fig. 23A–E View Figure 23 ). ALS have a small piriform field with tartipores ( Townley & Tillinghast, 2003; Fig. 23B, E, F View Figure 23 ). PMS have between one and two aciniform spigots between the cylindrical and the minor ampullate spigot ( Fig. 19D View Figure 19 ). PLS have between one and three aciniform spigots ( Fig. 19E View Figure 19 ); base of the peripheral cylindrical spigot at least twice the diameter of the base of the distal one ( Fig. 19E View Figure 19 ). The flagelliform and aggregate gland spigots (araneoid ‘triplet’) are well developed ( Fig. 19E View Figure 19 ). Triplet absent in the adult male (studied in L. thoracica ). Epiandrous fusules (studied in L. thoracica , Fig. 19F, G View Figure 19 ) arranged linearly in sockets containing from nine to two fusules; also a few individual fusules (without socket; Fig. 19F, G View Figure 19 ). Tracheal system haplotracheate [studied in L. thoracica (see Blest, 1976)].
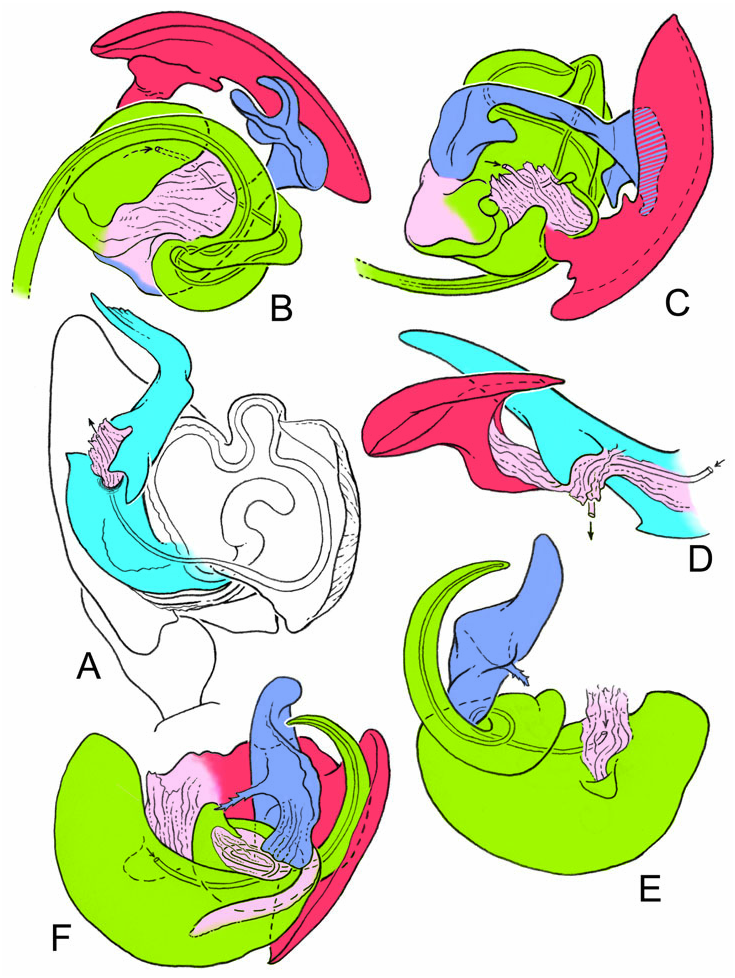
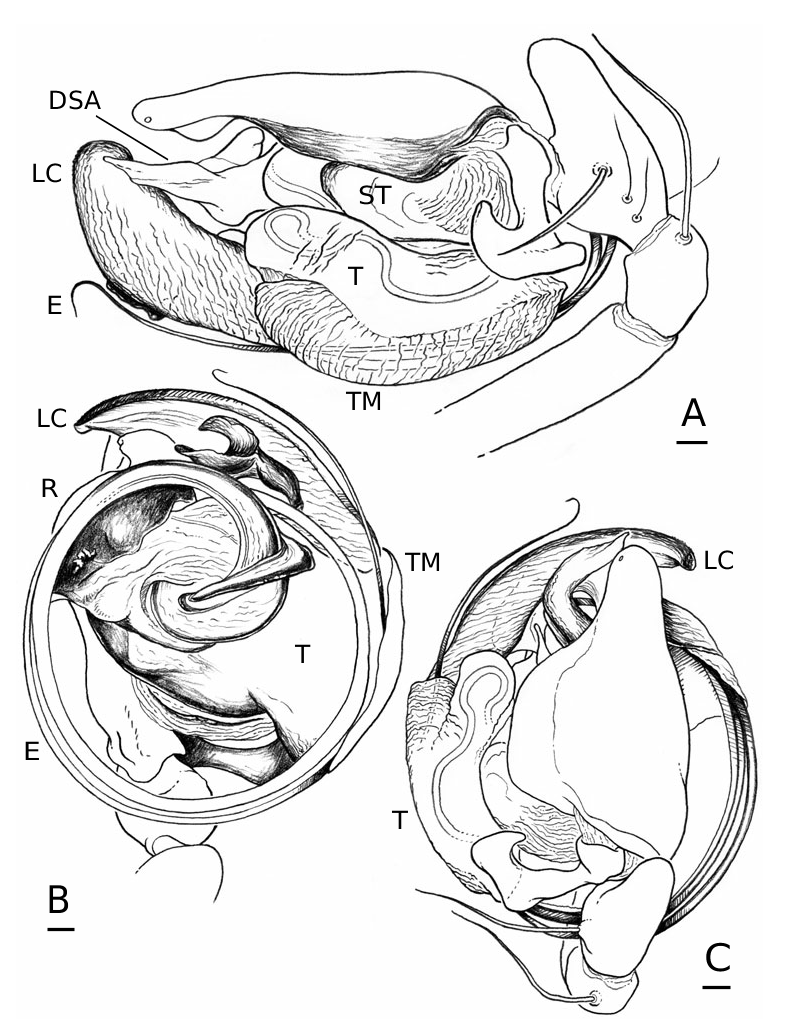
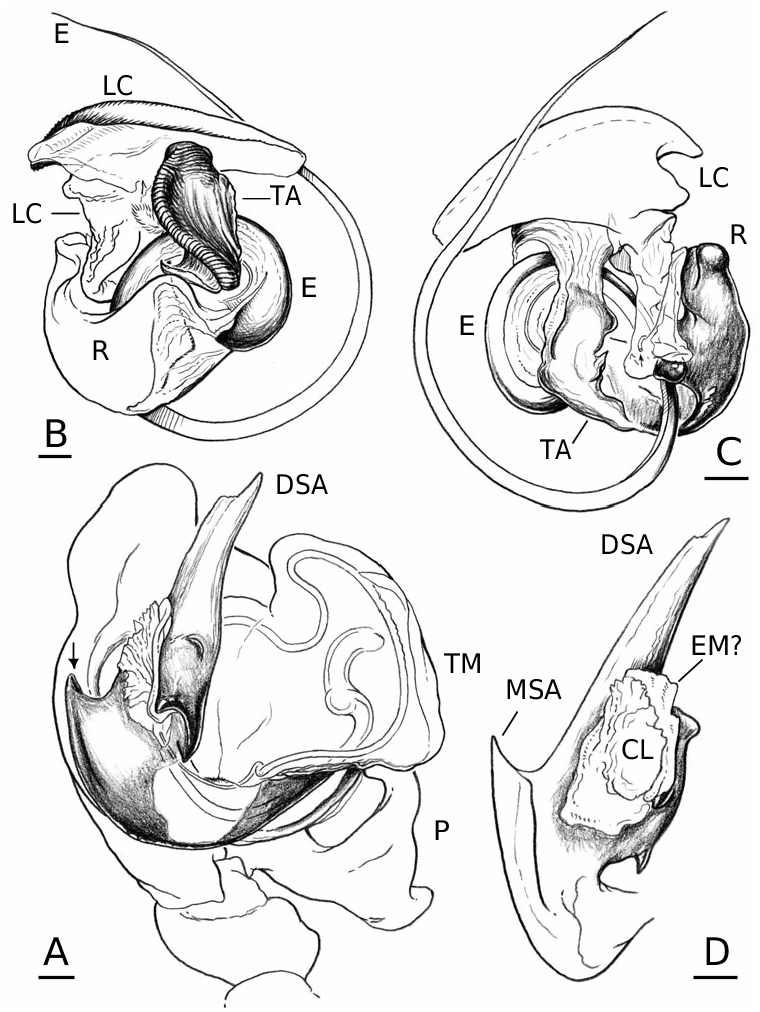
Male palpal tibia with dorsal process, one prolateral and two retrolateral trichobothria. Paracymbium with a caudad apophysis in the medial branch. Tegulum with an ectal tegular flap, membranous and running along its longitudinal axis. Suprategulum with a bifid suprategular apophysis adjacent to the column, a pointed apophysis, mesal to the suprategular foramen and a distal suprategular apophysis that is more or less flat in section. Typical embolic membrane absent. Lamella characteristica sheath-like, with a longitudinal fold housing the distal region of the embolus. Terminal apophysis morphology diverse. Embolus filiform, of varying length, coiling anticlockwise (left palp, ventral). Epigynum provided with anterior flaps on both sides of a ventral plate scape (without socket) and conspicuous posterior lateral lobes. Spermathecae Ushaped with anteriorly orientated fertilization ducts.
Composition
Three species: Labulla thoracica , L. flahaulti and L. machadoi sp. nov.
Phylogenetics
The monophyly of Labulla is supported by the following putative synapomorphies: marginal suprategular apophysis, ectal tegular flap, absence of embolic membrane, ectal position of the terminal apophysis, lamella sheath-like with a longitudinal fold housing the embolus, anterior epigynal flaps, and large posterior lateral epigynal lobes (see Discussion).
Natural history
Most of what we know about Labulla is based on the central European species Labulla thoracica . Information about the natural history of this species is given under the species description.
Distribution
Europe.
Misplaced species
The following three species do not belong in the genus Labulla :
‘ Labulla View in CoL ’ impudica Denis, 1945 . See comments and description under Pecado View in CoL gen. nov.
‘ Lepthyphantes View in CoL ’ insularis Saito, 1935 . This species was described based on a single female specimen from Sakhalin Island (located between the Sea of Japan and the Sea of Okhotsk). Tanasevitch & Eskov (1987: 194) remarked that Lepthyphantes insularis did not belong in Lepthyphantes View in CoL , based on Saito’s (1935, fig. 1b) epigynum illustration. Tanasevitch & Eskov did not provide any new illustrations or redescription of insularis , nor did they examine the type. In their view, Lepthyphantes insularis should probably be transferred to Labulla View in CoL , and it could be a (junior) synonym of L. thoracica View in CoL , although they did not formalize any transfer or new synonymy. Marusik et al. (1993: 75) synonymized L. insularis with Labulla chikunii Oi, 1979 [now Weintrauboa chikunii (Oi) , in the family Pimoidae View in CoL ], but this unjustified synonymy was rejected by Hormiga (2003: 276). Unfortunately, Tanasevitch & Eskov’s (1987) casual and perfunctory remarks on the affinities of L. insularis with Labulla thoracica View in CoL have been formalized in Platnick’s (2004) catalogue, where insularis appears as a valid species of Labulla View in CoL . We formally reject here the placement of Lepthyphantes insularis in the genus Labulla View in CoL , based on the absence of the two synapomorphies that we can assess in the external epigynum morphology of Saito’s illustration ( Saito, 1935, fig. 1b): insularis lacks both the anterior epigynal flaps and posterior epigynal lateral lobes that characterize Labulla View in CoL as defined in the present work. Additional support for this conjecture is provided by the lack of pigmentation around the PMEs (compare Saito’s fig. 1a with our Fig. 1 View Figure 1 ) and the almost uniform dark brown coloration of the prosoma of insularis (unlike any Labulla species ). But as Tanasevitch & Eskov (1987: 194) have also remarked (and with which we agree), insularis is not congeneric with the type species of Lepthyphantes View in CoL [ L. minutus (Blackwall, 1833) View in CoL ]. This is hardly surprising if we examine Saito’s (1935: 59) argument for placing insularis in the genus Lepthyphantes View in CoL : ‘The spider apparently resembles the European species, L. minutus View in CoL in general appearance and coloration, but it is distinct in having an epigynum which has only a tongue-like scape and lacks the parmula, while L. minutus View in CoL has both well defined.’ Saito’s type material, formerly at the University of Hokkaido, is presumably lost (H. Ono, in litt.). Because the species description is inadequate and insufficient to place insularis within another generic circumscription, we suggest that this species is cataloged as incertae sedis.
‘ Labulla View in CoL ’ nepula Tikader, 1970 . This species, described from Sikkim (located in India, between Nepal and Bhutan), was placed in Labulla View in CoL because it ‘resembles Labulla thoracica (Wider) View in CoL ’ ( Tikader, 1970: 21). How nepula resembles thoracica View in CoL , as opposed to many other linyphiids, is a mystery that Tikader does not explain in his publication. The type material (holotype female, and one male and one female paratypes) is presumably deposited in the National Zoological Collections, Zoological Survey of India (Calcutta), but we have not been able to borrow these specimens. Tikader’s description provides no information on genitalic morphology, other than his single illustrations of the palp and the epigynum. The palp (fig. 13d, presumably the left one) is illustrated in an unconventional view (at least for the single illustration of a new species), as it seems to be a dorsomesal view (e.g. no paracymbium can be discerned and in the embolic division, only the distal region of the embolus can be seen in some detail). None of the male Labulla View in CoL synapomorphies can be found on this figure. The epigynum (a ventral view, fig. 13c) does not reveal any of the generic synapomorphies either. Consequently, we reject the placement of nepula in the genus Labulla View in CoL . Because the species description is inadequate and insufficient to place nepula within another generic circumscription, we suggest that this species is catalogued as incertae sedis.
LABULLA THORACICA ( WIDER, 1834) View in CoL
( FIGS 1A, B View Figure 1 , 2 View Figure 2 , 3A–D View Figure 3 , 4A–C View Figure 4 , 5–7 View Figure 5 View Figure 6 View Figure 7 , 16–19 View Figure 16 View Figure 18 View Figure 19 , 20A, B View Figure 20 , 22E View Figure 22 )
Linyphia thoracica Wider, 1834: 254 , pl. 17, f. 180.
Linyphia cauta Blackwall, 1841: 655 . – Blackwall, 1864: 220, pl. 15, fig. 145.
Bathyphantes thoracicus Förster & Bertkau, 1883: 253 .
Labulla thoracica ( Wider, 1834) View in CoL : Simon, 1884: 262. – Chyzer & Kulczynski, 1894: 61, pl. 2, fig. 30. – Becker, 1896: 35, pl. 5, fig. 2. – Bösenberg, 1901: 92, pl. 8, fig. 111. – Simon, 1929: 625, 741, figs 942, 943. – Wiehle, 1952: 231, figs 9, 10. – Locket & Millidge, 1953: 375, fig. 224D,E. – Wiehle, 1956: 293, figs 481–485. – Merrett, 1963: 377, fig. 33A–E. – Miller, 1971: 222, pl. XLIII, fig. 4. – Palmgren, 1975: 77, figs 7.28–32. – Roberts, 1987: 142, figs 73b, 74a. – Heimer & Nentwig, 1991: 176, fig. 471. – Millidge, 1993: 146, fig. 1. – Roberts, 1995: 352. – Roberts, 1998: 364.
Diagnosis
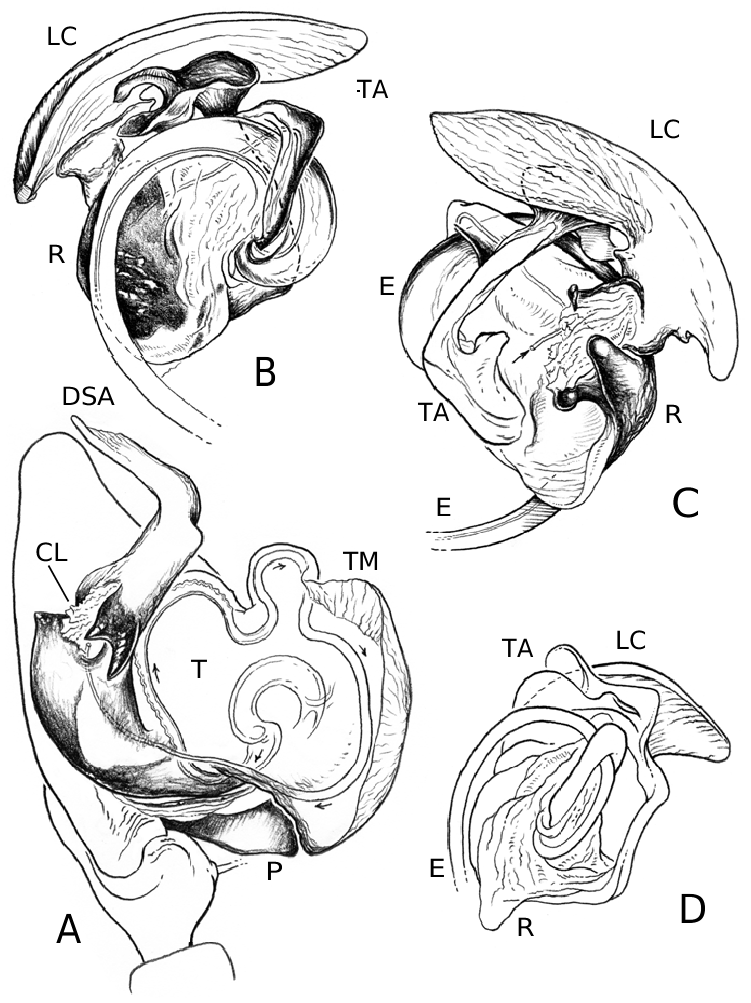
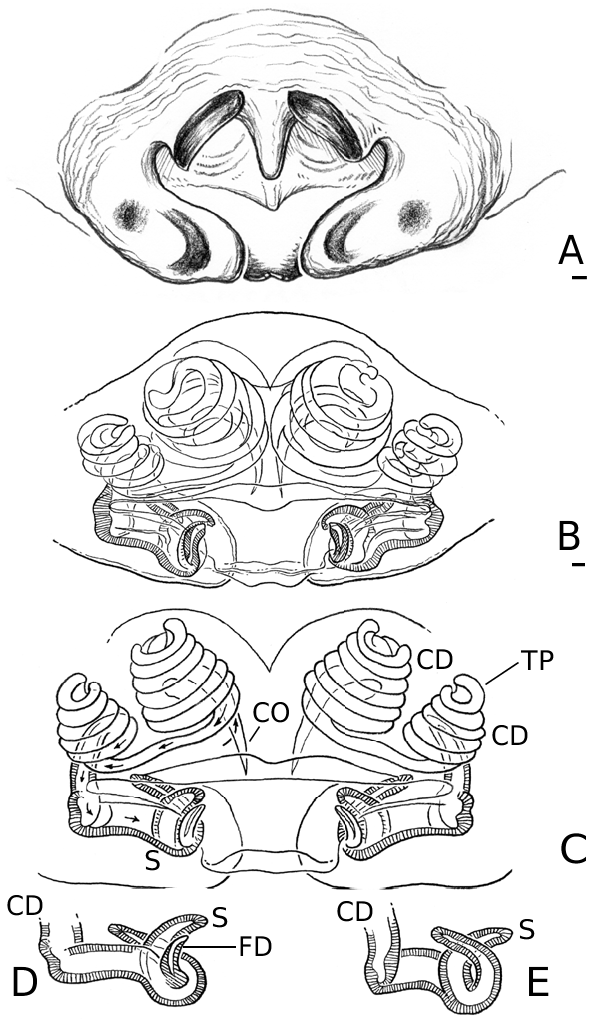
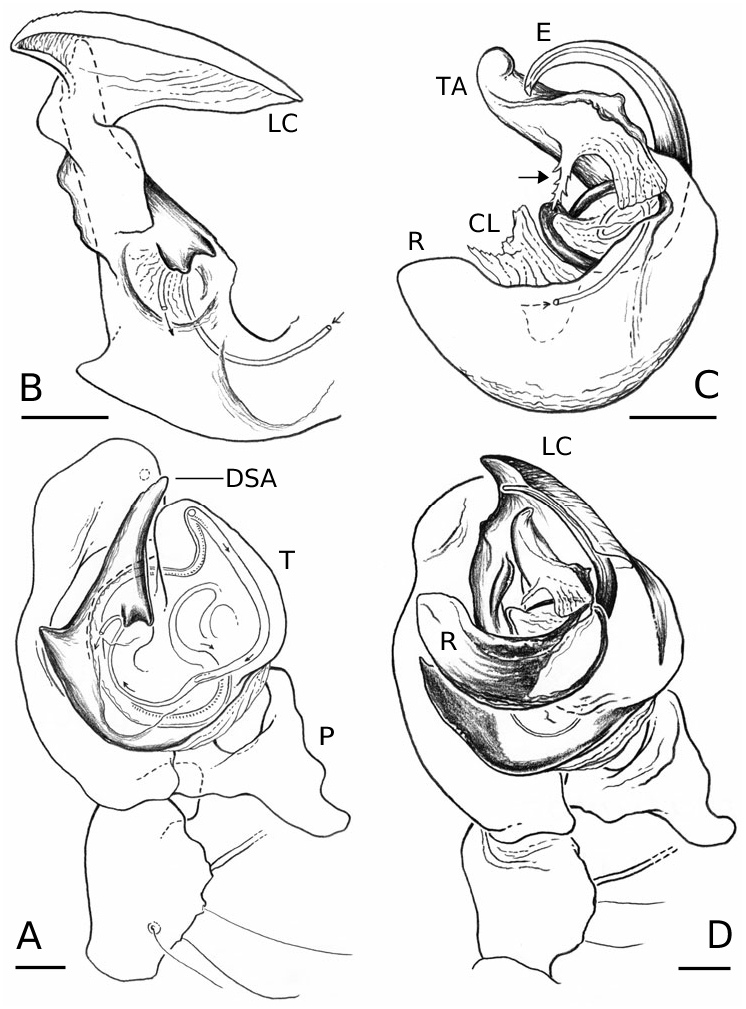
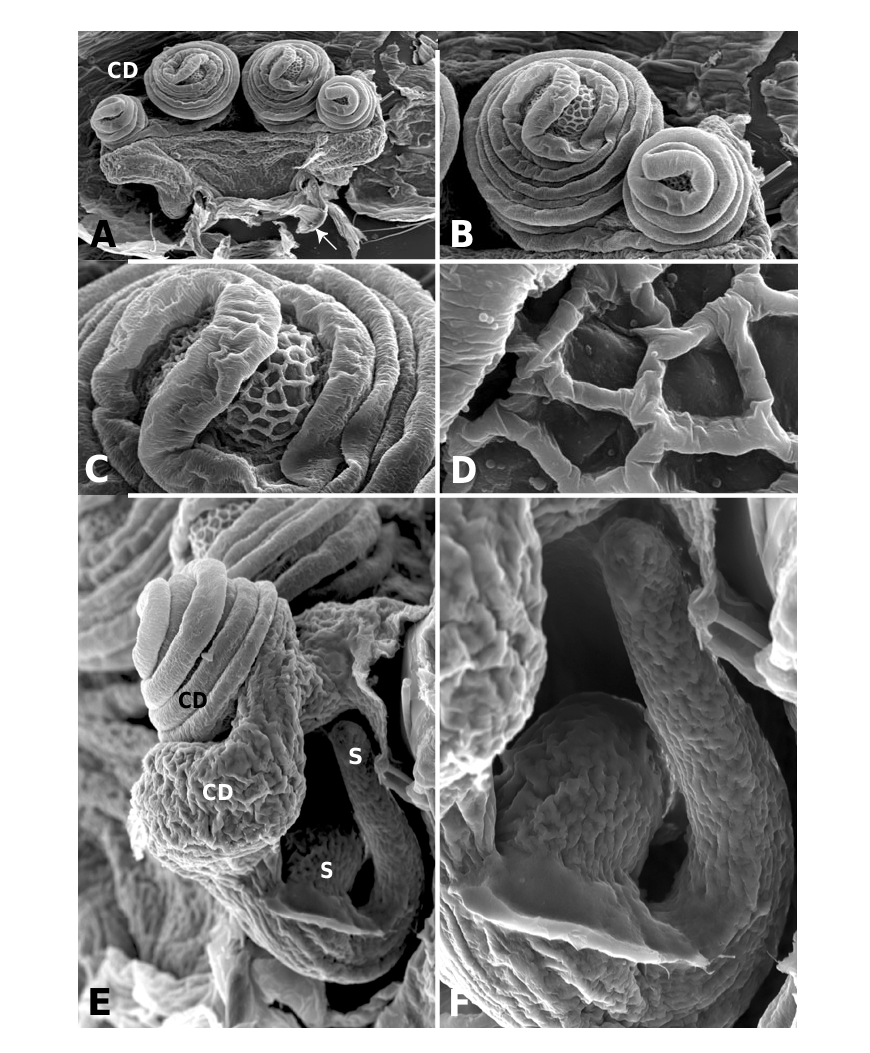
Males of L. thoracica can most easily be distinguished from other Labulla species by the presence of a long tegular flap ( Fig. 5A View Figure 5 , longer than in machadoi sp. nov.), long embolus (more than two complete turns, Fig. 5B View Figure 5 ), the shape of the terminal apophysis and the absence on it of a plumose process ( Fig. 5B View Figure 5 ), and the shape of the dorsal tibial apophysis (vertical and inclined over the cymbium, Fig. 5A View Figure 5 ). Females are diagnosed externally by the blade-like epigynal flaps ( Figs 3D View Figure 3 , 7A View Figure 7 ) and internally by the presence of two pairs of copulatory duct coils ( Fig. 7B View Figure 7 ).
Description
Male (from Denmark: Hestehaven, Rønde, 22 km north-east of Århus, 56∞17.46¢N, 10∞28.50¢E, 31.viii.1994, Bjørn, Christensen, Coddington, Griswold, Hormiga, Krat, Langemark, Scharff & Sørensen leg.; cat. no. ZMUC00004683). Total length 5.20. Cephalothorax 2.24 long, 2.00 wide. Sternum 1.12 long, 1.25 wide. Abdomen 3.04 long, 1.92 wide. Cephalothorax and chelicerae pale yellowish brown, with greyish brown borders and darker yellowish brown cephalic area ( Fig. 1A View Figure 1 ). The dark coloration extends from cephalic area to posterior rim of cephalothorax (midline). Black rings around all eyes ( Fig. 1A View Figure 1 ). Sternum dark greyish brown. Legs yellowish brown, with dark annulations. Abdomen light grey, with dark grey pattern and white spots ( Fig. 1A View Figure 1 ). Ventrum with three white dots in front of spinnerets. Cephalothorax pearshaped. Cheliceral stridulatory striae present, but faint. AME diameter 0.12. Clypeus height 5.75 times one AME diameter (raised cephalon). Chelicerae with three large widely spaced prolateral teeth and three small retrolateral (four in other specimen) more closely spaced proximal teeth. Femur I 4.40 long, 1.96 times the length of cephalothorax. Metatarsus I trichobothrium absent (see comments under ‘Variation’). Pedipalp as in Figures 4–6 View Figure 4 View Figure 5 View Figure 6 , 16 View Figure 16 , 17. Pedipalpal tibia with three trichobothria (one prolateral and two retrolateral) and a strong retrolateral spine. Epiandrous fusules (studied in two specimens) arranged more or less linearly in sockets, which contain from nine to two fusules. A few individual fusules, not in sockets, can also be found ( Fig. 19F, G View Figure 19 ).
Female (from same locality, and with same data as male). Total length 5.98. Cephalothorax 2.41 long, 1.89 wide. Sternum as long as wide (1.16). Abdomen 4.03 long, 2.60 wide. Same coloration as male, but with more pronounced annuli on all legs and pedipalps ( Figs 1B View Figure 1 , 2A View Figure 2 ). Cephalothorax pear-shaped. Cheliceral stridulatory striae present ( Fig. 19A View Figure 19 ). AME diameter 0.18. Clypeus height 2.04 times one AME diameter. Chelicerae ( Fig. 22E View Figure 22 ) with three large widely spaced prolateral teeth and three to four small retrolateral closely spaced proximal teeth (three on one chelicera and four on the other). Femur I 3.20 long, 1.33 times the length of cephalothorax. Metatarsus I trichobothrium 0.21. Epigynum as in Figures 3D View Figure 3 , 7 View Figure 7 , 18 View Figure 18 , 20A, B View Figure 20 . Spinnerets ( Fig. 19B–E View Figure 19 ) typical for the family, with two aciniform spigots on PMS ( Fig. 19D View Figure 19 ) and three on PLS ( Fig. 19E View Figure 19 ).
Variation
Male total length ranges from 4.00 to 5.20 (N = 5; average 4.64). Female total length ranges from 4.32 to 6.20 (N = 6; average 5.14). Male cephalothorax ranges in length from 2.00 to 2.32 (N = 5; average 2.18). Female cephalothorax ranges in length from 1.84 to 2.41 (N = 6; average 2.17). Male femur I ranges in length from 3.60 to 4.40 (N = 5; average 4.06). Female femur I ranges in length from 2.93 to 3.52 (N = 6; average 3.10). Trichobothrium metatarsus I varies between 0.21 0.30 and is occasionally absent in males ( Roberts, 1987).
Distribution
Labulla thoracica appears in faunal listings of most Central and Northern European countries, including the British Isles and Ireland, France, Italy, Belgium, the Netherlands, Liechtenstein, Switzerland, Austria, Germany, Poland, Czech Republic, Croatia, Slovenia, Slovakia, Yugoslavia, Romania, Denmark, Sweden, Norway, Finland, Ukraine and Russia. It has never been recorded from Spain or Portugal.
Natural history
Numerous works on European spiders provide general information about the natural history of this species (e.g. Simon, 1884; Nielsen, 1928; Locket & Millidge, 1953; Wiehle, 1956; Bristowe, 1958; Roberts, 1987; Harvey, Nellist & Telfer, 2002). According to these accounts Labulla thoracica is relatively common in the British Isles and Central Europe, spinning sheet webs, mainly at ground level, in wooded areas. The webs are usually found at the base of trees between the buttresses, in overhanging banks, under logs and in a variety of shady habitats, including man-made structures such as cellars and outhouses. In the British Isles adults of both sexes are found mainly between August and October, but females and occasionally males can be found in most months ( Harvey et al., 2002). In Hestehaven ( Denmark), adults were found from May to December ( Scharff et al., 2003). Eggsacs are concealed under loose objects (bark, moss, etc.) on the ground some distance from the web ( Nielsen, 1928; Bristowe, 1958: fig. 423). Egg sacs are slightly domed, covered with white curled silk and approximately 1 cm in diameter. Eggs are light yellow ( Nielsen, 1928).
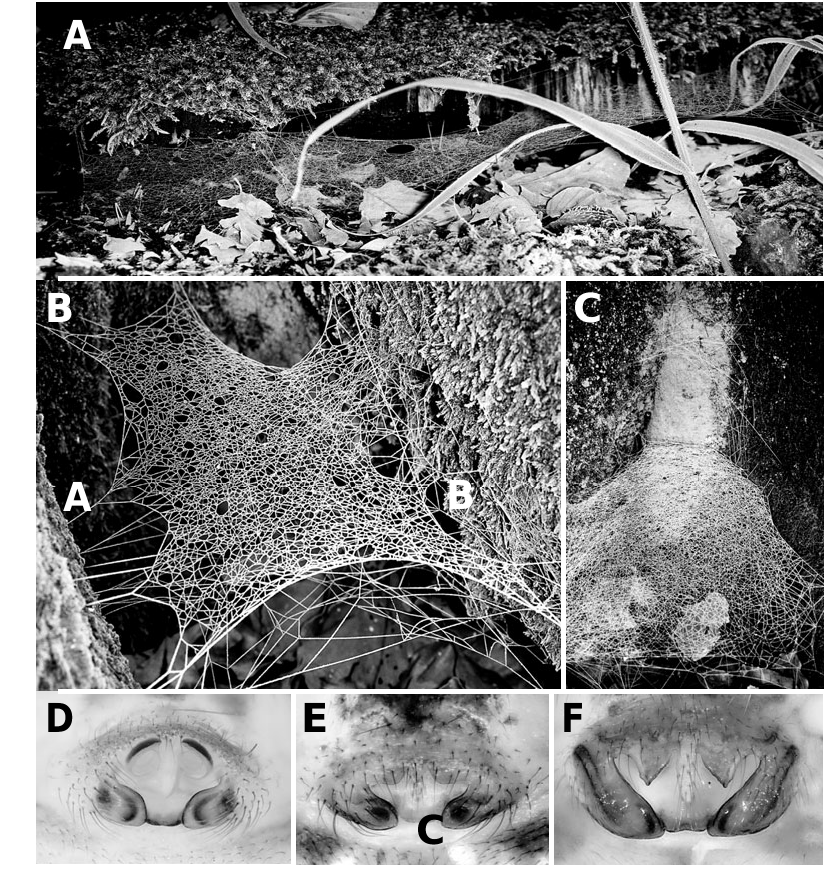
We have collected this species throughout Denmark (e.g. Scharff et al., 2003), where it is abundant and common in wooded areas. We have photographed seven different webs (two male and five female adult webs), and four of them are depicted in Figures 2B–D View Figure 2 , 3A–C View Figure 3 to illustrate the general architecture of this species. During the daytime the spiders hide in a retreat in the less exposed area of the web and they are rarely seen; at night they are found, upside-down (as in all linyphiids), in the centre of the web. Adult males are often found together with females, suggesting that when males move into female webs they occupy their webs for some time after copulation.
Webs consist of a main platform or sheet, with the perimeter often delimited by relatively long silk lines in the same plane (e.g. Figs 2B View Figure 2 , 3B View Figure 3 ) as is generally the case in non-erigonine linyphiids. The mesh of the main platform shows the usual pattern of linyphiid mesh ( Fig. 2D View Figure 2 ), with silk lines that can be ‘traced’ for quite some length. The upper scaffolding is most often absent, except perhaps for a few vertical lines, but some webs do show some upper mesh ( Fig. 3A View Figure 3 ). Presence or absence of any upper scaffolding is most likely determined in part by the availability of attachment points, but this architectural feature, although variable, is generally absent. By contrast, a three-dimensional mesh of silk lines is usually found under the main platform ( Fig. 2C View Figure 2 ), although in some webs this under-mesh is lacking altogether. We have found a rather different pattern in the main platform silk mesh of male ( Fig. 3B View Figure 3 ) and female webs ( Figs 2B, D View Figure 2 , 3C View Figure 3 ), as well as in the dimensions of the platform (adult male webs seem smaller); but we lack sufficient information to report whether these differences are consistent. It is also unknown for how long males remain in their webs after reaching maturity (the PLS triplet is reduced to nubbins in the adult male). As is generally the case for large linyphiid webs, they are maintained for long periods of time and often show signs of repair and senescence. Nielsen (1928) suggests that the web of Labulla traps small flying insects (such as fungus gnats) that seek shelter in the dark cavities of tree buttresses. Clouds of small flies emerge from these cavities, when one tries to catch Labulla thoracica on the webs in between the tree buttresses ( Nielsen, 1928; our own observations).
When disturbed these animals often ‘drop dead’ to the ground, as is commonly seen in other linyphiids (e.g. in Orsonwelles species , Hormiga, 2002: fig. 4F).
Mating is in type 2 position, as is typical of Linyphiidae [see Foelix (1996) for definitions and additional references]. Gerhardt (1921, 1923, 1925, 1928) studied the mating behaviour of this species [see Huber (1998) for a summary of Gerhardt’s observations].
Material examined
Germany: Rothiemurchus Forest , 12.ix.1965, J.A. L.C., 1 male ( AMNH) ; Denmark: Zealand , Vesterlyng, south-west of Havnsø, 55∞44.6¢N, 11∞17.5¢E, 16.viii.2003, 1 female, G. Hormiga & N. Scharff leg. ( ZMUC) ; Zealand , Dragerup Forest near Tjebberup, 55∞42.8¢N, 11∞47.5¢E, 15.viii.2003, 1 male & 1 female, G. Hormiga & N. Scharff leg. ( ZMUC) ; same locality and data, 2 males & 1 female, G. Hormiga & N. Scharff leg. ( ZMUC) ; Jutland , Hestehaven , Rønde, 22 km north-east of Århus, 56∞17.46¢N, 10∞28.50¢E, 30.viii.1994, 2 males & 2 females, Bjørn, Christiansen, Coddington, Griswold, Hormiga, Krat, Langemark, Scharff & Sørensen leg. ( ZMUC00004682 View Materials ) ; same locality and same data, 12 females & 10 males ( ZMUC00004687 View Materials ) ; same locality, same data, 3 females & 4 males ( ZMUC) ; Zealand , Roden and Frejlev Forest, 7–13.ix.1883, 1 female, Løvendal leg. ( ZMUC00004691 View Materials ) ; Jutland , Lille Vildmose, Høstemark Forest, 56∞56¢N, 10∞13¢E, 20–24.viii.1999, 9 males & 6 females, N. Scharff leg. ( ZMUC00007706 View Materials ) .
LABULLA FLAHAULTI SIMON, 1914 View in CoL
( FIGS 1C, D View Figure 1 , 3E View Figure 3 , 4D–F View Figure 4 , 8–10 View Figure 8 View Figure 9 View Figure 10 , 20C, D View Figure 20 , 22F View Figure 22 )
Labulla flahaulti Simon, 1914: 478 View in CoL (syntype material in MNHN, examined); Simon, 1929: 626 and 741, figs 944, 945; Fage, 1935: 179, fig. 2D.
Diagnosis
Males of L. flahaulti can most easily be distinguished from other Labulla species by the smaller size of the palps ( Fig. 1C View Figure 1 ), the short length of the embolus (less than half a coil, Fig. 9C View Figure 9 ), the large size of the paracymbial process ( Fig. 8A, B View Figure 8 ), the shape of the terminal apophysis, with a small plumose process on it ( Fig. 9C View Figure 9 – arrow) and the shape of the dorsal tibial apophysis (blunt, orientated ectally, Fig. 8A View Figure 8 ). Females are diagnosed externally by the subtle epigynal flaps ( Figs 3E View Figure 3 , 10A View Figure 10 ) and internally by the absence of any copulatory duct coils ( Fig. 10B View Figure 10 ).
Description
Male (from Spain: Gipuzkoa, San Sebastián, Igara, 50 m, 7.ix.2002, A. de Castro & A. Casis leg., AZE). Total length 4.45. Cephalothorax 2.10 long, 1.55 wide. Sternum as long as wide (1.11). Abdomen 2.73 long, 1.56 wide. Cephalothorax and chelicerae pale greyish white, with greyish brown borders and cephalic area ( Fig. 1C View Figure 1 ). The dark coloration extends from cephalic area to posterior rim of cephalothorax. Black rings around all eyes. Sternum dark greyish brown. Legs yellowish brown, with dark annulations. First two pairs of legs darker than the rest. Abdomen light grey, with dark grey pattern ( Fig. 1C View Figure 1 ). Ventrum with one to three patches of white dots. Two posterior to the booklungs, just behind the epigastric furrow, and one patch just in front of the spinnerets. Patch in front of spinnerets may include two to five smaller white dots. Cephalothorax pear-shaped. Cheliceral stridulatory striae present, but faint. AME diameter 0.14. Clypeus height 2.23 times one AME diameter. Chelicerae with three large widely spaced prolateral teeth and three small retrolateral (four in other specimen) more closely spaced proximal teeth. Femur I 4.00 long, 1.91 times the length of cephalothorax. Trichobothrium metatarsus I 0.24. Pedipalp as in Figures 8 View Figure 8 , 9 View Figure 9 , 21. Pedipalpal tibia with one prolateral and two retrolateral trichobothria.
Female (from same locality, and with same data as male). Total length 5.40. Cephalothorax 2.30 long, 1.88 wide. Sternum as long as wide (1.20). Abdomen 5.07 long, 2.54 wide. Cephalothorax and chelicerae pale yellowish brown, with greyish brown borders and cephalic area ( Fig. 1D View Figure 1 ). The dark coloration extends from cephalic area to posterior rim of cephalothorax. Black rings around all eyes. Sternum dark greyish brown. Legs yellowish brown, with dark rings (annulations). Abdomen light grey, with dark grey pattern ( Fig. 1D View Figure 1 ). Cephalothorax pear-shaped. Cheliceral stridulatory striae present. AME diameter 0.17. Clypeus height 1.91 times one AME diameter. Chelicerae ( Fig. 22F View Figure 22 ) with three large widely spaced prolateral teeth and four small retrolateral closely spaced proximal teeth. Femur I 4.05 long, 1.76 times the length of cephalothorax. Trichobothrium metatarsus I 0.20. Epigynum as in Figures 3E View Figure 3 , 10 View Figure 10 , 20C, D View Figure 20 .
Variation
Male total length ranges from 4.40 to 4.45 (N = 2; average 4.43). Female total length ranges from 4.80 to 6.00 (N = 5; average 5.59). Male cephalothorax ranges in length from 1.90 to 2.10 (N = 2; average 2.00). Female cephalothorax ranges in length from 2.25 to 2.60 (N = 5; average 2.40). Male femur I ranges in length from 3.90 to 4.00 (N = 2; average 3.95). Female femur I ranges in length from 3.12 to 4.05 (N = 6; average 3.44). The length of the epigynal scape is variable.
Distribution
Southern France and northern Spain ( Simon, 1914, 1929; Bosmans & Keer, 1985; Ribera & Hormiga, 1985; Bosmans, Maelfait & Kimpe, 1986; Ledoux, Emerit & Pinault, 1996; Castro, 2004).
Natural history
Simon (1914, 1929) reported that in France L. flahaulti lives in fir and beech forests and that their large webs, similar to those of L. thoracica , are often found in overhanging banks. Ribera & Hormiga (1985: 186) have reported this species in conifer and beech forests in northern Spain (c. 960–1415 m), with small numbers of adults collected in pitfall traps between July and September. Ledoux et al. (1996) have reported this species from Nohèdes (Pyrénées Orientales). Bosmans et al. (1986: 75) reported it from St. Lary (Midi Pyrénées) in July at 1500– 1700 m (montane zone), in fir forest ( Picea abies ). Castro (2004) provides the most detailed account available on the natural history of Labulla flahaulti , based on his ecological work on the Cantabrian oak forests of the Basque country. In this region flahaulti is mainly found in forested areas, where their webs can be seen on tree trunks. Males are adult in September, females between September and November and juveniles are found between spring and autumn. This suggests that mating takes place at the end of the summer or early in autumn and that the species over winters in the eggsac. Although Castro (2004) collected this species in almost all forest strata, flahaulti seems to prefer the higher parts (tree trunks, branches, etc.).
Material examined
Spain: Gipuzkoa , San Sebastián, Igara, 50 m, 7.ix.2002, 8 females & 2 males (incl. those described above), A. de Castro & A. Casis leg. ( AZE) ; Huesca , San Juan de la Peña, C. Pedrocchi, 1 male, Pinar medio, 29.viii.1977 ; Abetal Alto , 1 male, 16.viii.1977 ( UB) . France: Fôret de la Massane de Valmanya du Canigou , 3 males, 8 females, 1 juv., 22873 (syntype series, in part; MNHN) .
LABULLA MACHADOI SP. NOV.
( FIGS 1E, F View Figure 1 , 3F View Figure 3 , 11– 13 View Figure 11 View Figure 12 View Figure 13 , 20E, F View Figure 20 , 22A–D,G View Figure 22 , 23 View Figure 23 )
Type. Male holotype and female paratype from Hto. Dias Ferreira, Porto, Portugal; March 1937, A.B. Machado leg. (cat. no. 267; deposited in MB) .
Etymology
Named after the collector, António de Barros Machado.
Diagnosis
Males of L. machadoi sp. nov. can most easily be distinguished from other Labulla species by the presence of a long tegular flap ( Fig. 11A View Figure 11 , shorter than in thoracica ), long embolus (only one complete turn, Fig. 11B View Figure 11 ), the hooked paracymbial process ( Fig. 11A– C View Figure 11 ), the shape of the terminal apophysis, with a plumose process on it ( Fig. 11B View Figure 11 ), and the shape of the dorsal tibial apophysis (pointed, orientated ectally, Fig. 11A, C View Figure 11 ). Females are diagnosed externally by the V-shaped epigynal flaps ( Figs 3F View Figure 3 , 13A View Figure 13 ) and internally by the presence of one pair of copulatory duct coils ( Fig. 13C View Figure 13 ).
Description
Male holotype: Total length 4.20. Cephalothorax 2.10 long, 1.70 wide. Sternum 1.20 long, 1.12 wide. Abdomen 2.40 long, 1.60 wide. Cephalothorax and chelicerae yellowish brown, darker in cephalic area. Black rings around all eyes ( Fig. 1E View Figure 1 ). Sternum greyish brown. Legs same colour as cephalothorax, but with dark annulations. Abdomen light grey, with dark grey pattern ( Fig. 1E View Figure 1 ). Shape of cephalothorax more oval than in L. thoracica and L. flahaulti ( Fig. 1A, C, E View Figure 1 ). Cheliceral stridulatory striae present. AME diameter 0.16. Clypeus height 2.52 times one AME diameter. Chelicerae with three large widely spaced prolateral teeth and four small retrolateral closely spaced proximal teeth. Femur I 3.44 long, 1.64 times the length of cephalothorax. Trichobothrium metatarsus I 0.22. Pedipalp as in Figures 11 View Figure 11 , 12 View Figure 12 , 22A–D View Figure 22 . Pedipalpal tibia with one prolateral and two retrolateral trichobothria.
Female paratype (together with male holotype): Total length 5.60. Cephalothorax 2.20 long, 1.90 wide. Sternum as long as wide (1.15). Abdomen 3.90 long, 2.60 wide. Same colour pattern as male, but brown instead of yellowish brown ( Fig. 1F View Figure 1 ). Cephalothorax pearshaped. Cheliceral stridulatory striae absent. AME diameter 0.14. Clypeus height 2.50 times one AME diameter. Cheliceral dentation as in male. Femur I 2.80 long, 1.27 times the length of cephalothorax. Trichobothrium metatarsus I 0.22. Epigynum as in Figures 3F View Figure 3 , 13 View Figure 13 , 20E View Figure 20 , F. Spinnerets as in Figure 23 View Figure 23 . PMS and PLS with only one aciniform spigot .
Variation
Male total length ranges from 4.20 to 4.80 (N = 4; average 4.42). Female total length ranges from 3.36 to 5.60 (N = 7; average 4.28). Male cephalothorax ranges in length from 2.10 to 2.40 (N = 4; average 2.26). Female cephalothorax ranges in length from 1.60 to 2.20 (N = 7; average 1.90). Male femur I ranges in length from 3.44 to 4.16 (N = 4; average 3.85). Female femur I ranges in length from 2.12 to 3.12 (N = 7; average 2.59). The shape of the epigynal flaps and the length of the scape is variable.
Distribution
Only known from Portugal.
Natural history
Specimens have been taken under moss in a ‘Horto’ (garden/orchard) in Porto. Adult males present in October and December, and adult females in March, August and October.
Material examined
Portugal: Porto, Hto Dias Ferreira , 31.x.1937, 4 females and 1 male together with holotype and paratype, A.B. Machado leg. (cat. no. 267; MB) ; same locality, iii.1937, 5 females, A.B. Machado leg. (cat. no. 3; MB) ; Amarante , iii.1937, 1 female, A.B. Machado leg. (cat. no. 276; MB) ; Joane , 3.x.1941, 2 females and 3 juveniles, A.B. Machado leg. (cat. no. 983; MB) ; Joane , 10.x.1937, 1 female, A.B. Machado leg. (cat. no. 248; MB) ; Joane, Baltar , 7.x.1937, 5 females, A.B. Machado (cat. no. 225; MB) ; Paredes de Coura, Mantelcus ( Mate ), 12.x.1940, 2 females, A.B. Machado leg. (cat. no. 677; MB) ; Paredes de Coura, Ramalhosa de Vascões , 25.viii.1940, 1 female, A.B. Machado leg (cat. no. 662; MB) ; Joane, Ciridade , 4.x.1937, 1 male, A.B. Machado (cat. no. 224; MB) ; Porto, 5.xii.1937, 1 male, A.B. Machado leg. (cat. no. 297; MB) .

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Labulla
| Hormiga, Gustavo & Scharff, Nikolaj 2005 |
Labulla flahaulti
| Fage L 1935: 179 |
| Simon E 1929: 626 |
| Simon E 1914: 478 |
Labulla
| Simon E 1894: 707 |
| Simon E 1884: ) |
| Wider 1834: 254 |
Labulla thoracica ( Wider, 1834 )
| Roberts MJ 1998: 364 |
| Roberts MJ 1995: 352 |
| Millidge AF 1993: 146 |
| Heimer S & Nentwig W 1991: 176 |
| Roberts MJ 1987: 142 |
| Palmgren P 1975: 77 |
| Miller F 1971: 222 |
| Merrett P 1963: 377 |
| Wiehle H 1956: 293 |
| Locket GH & Millidge AF 1953: 375 |
| Wiehle H 1952: 231 |
| Simon E 1929: 625 |
| Bosenberg W 1901: 92 |
| Becker L 1896: 35 |
| Chyzer C & Kulczynski W 1894: 61 |
| Simon E 1884: 262 |
Bathyphantes thoracicus Förster & Bertkau, 1883: 253
| Forster A & Bertkau P 1883: 253 |
Linyphia cauta
| Blackwall J 1864: 220 |
| Blackwall J 1841: 655 |
Linyphia thoracica
| Wider 1834: 254 |