
Blepharisma penardi, Yan & Fan & Chen & Li & Warren & Al-Farraj & Song, 2016
|
publication ID |
https://doi.org/10.1111/zoj.12369 |
|
persistent identifier |
https://treatment.plazi.org/id/DA7BBB25-1F23-A13D-FC80-031F497BF9A6 |
|
treatment provided by |
Marcus |
|
scientific name |
Blepharisma penardi |
| status |
|
COMMENTS ON BLEPHARISMA UNDULANS STEIN, 1867 View in CoL
This well-known organism has been reported on numerous occasions from sites all over the world and thus its identity is relatively easy to establish ( Stein, 1867; Kahl, 1932; Suzuki, 1954; Bhandary, 1962; Cela, 1972; Dragesco & Dragesco-Kernéis, 1991; Lee & Shin, 2009). The Qingdao population corresponds closely with both the original report ( Stein, 1867) and the redescription by Kahl (1932) in terms of its bright pink coloration and the pinkish cortical granules, general morphology, pattern of ciliature, possession of two macronuclear nodules, and habitat ( Fig. 6 View Figure 6 ; Table 4).
Compared with the South Korean and Argentinian populations ( Cela, 1972; Lee & Shin, 2009), the Qingdao population has only one dissimilar feature, i.e. the body shape: the Qingdao population is stably lanceolate or elongate-pyriform with the anterior end pointed and posterior end widely rounded (vs. spindle- or batshaped with the posterior third slightly narrowed in the South Korean and Argentinian populations; Fig. 6F, H View Figure 6 ). It has long been known that body shape in Blepharisma undulans is variable and largely dependent upon nutrition, with well-nourished cells usually assuming a pyriform shape, expanded posteriorly, and underfed organisms having an elongate spindle shape ( Giese, 1973). We therefore believe that the difference in body shape between the Qingdao population of B. undulans and the South Korean and Argentinian populations to be population- or environment-dependent and not significant for taxonomy. The conservative nature of B. undulans is also supported by the molecular data, the SSU rDNA sequence obtained in this study being identical to that provided by Schmidt et al. (2007).
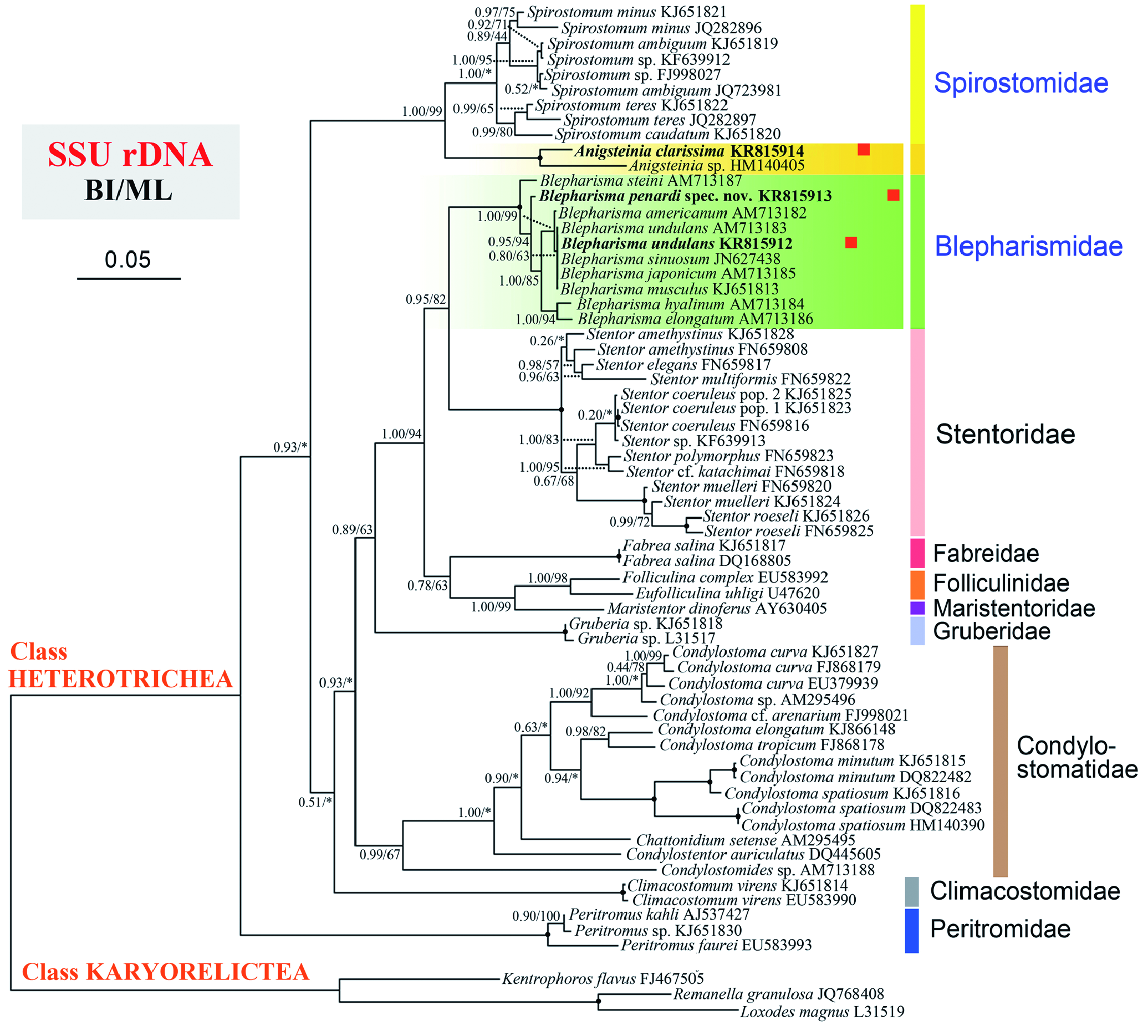
PHYLOGENETIC ANALYSES
The overall topology of our phylogenetic tree is congruent with that of Shazib et al. (2014). The newly sequenced species A. clarissima clusters with its congener, Anigsteinia sp. ( HM140405 View Materials ), on a branch that is deeply divergent from Blepharisma , thus supporting the morphological evidence for separating Anigsteinia from Blepharisma ( Isquith, 1968) . The two Anigsteinia spp. form a clade that is sister to the Spirostomum assemblage and so the family Spirostomidae is monophyletic with Anigsteinia included. Therefore, the molecular data also support the morphological evidence, i.e. the uniquely posteriorly thickened paroral membrane and the terminal contractile vacuole system with a long canal, for the assignment of the former to the family Spirostomidae , as suggested previously ( Isquith, 1968; Isquith & Repak, 1974; Shazib et al., 2014).
The addition of two new sequences of Blepharisma verifies the monophyly of this genus. As shown in Figure 7 View Figure 7 , B. penardi sp. nov. occupies a separate branch within the Blepharisma clade and is clearly separat- ed from its congeners (including the morphologically similar species, B. steini ), which supports our morphological evidence for its establishment as a new species. Furthermore, apart from the difference in sequence length (the Chinese population is 47 bp shorter at the 5′ end and 33 bp longer at the 3′ end), the SSU rDNA sequence of the Chinese population of B. undulans is identical to that of the German population ( AM713183 View Materials ) investigated by Schmidt et al. (2007). It is noteworthy, however, that phylogenetic relationships within the genus Blepharisma are largely unresolved because the SSU rDNA sequences of several species are either identical or differ only negligibly ( Fernandes et al., 2013). Consequently, sequence data of genes that evolve more rapidly than SSU rDNA, e.g. the mitochondrial cytochrome oxidase I gene or the ribosomal internal transcribed spacer regions (ITS1 and IT2), are needed in order to resolve this problem.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.