
Proszynellus, Patoleta, Barbara & Żabka, Marek, 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3926.2.6 |
|
publication LSID |
lsid:zoobank.org:pub:3367EA0E-C55F-4AB1-A88C-BF8830BA7DFA |
|
DOI |
https://doi.org/10.5281/zenodo.6097710 |
|
persistent identifier |
https://treatment.plazi.org/id/DE3A1527-E970-FFE8-E3D5-FF65FA8F7D1D |
|
treatment provided by |
Plazi |
|
scientific name |
Proszynellus |
| status |
gen. nov. |
Proszynellus View in CoL gen. nov.
Type species: Proszynellus nasalis sp. nov.
Etymology. The genus is described in honour of Professor Jerzy Prószyński, world authority in salticid taxonomy and biogeography, on his 80th anniversary. The name is masculine in gender.
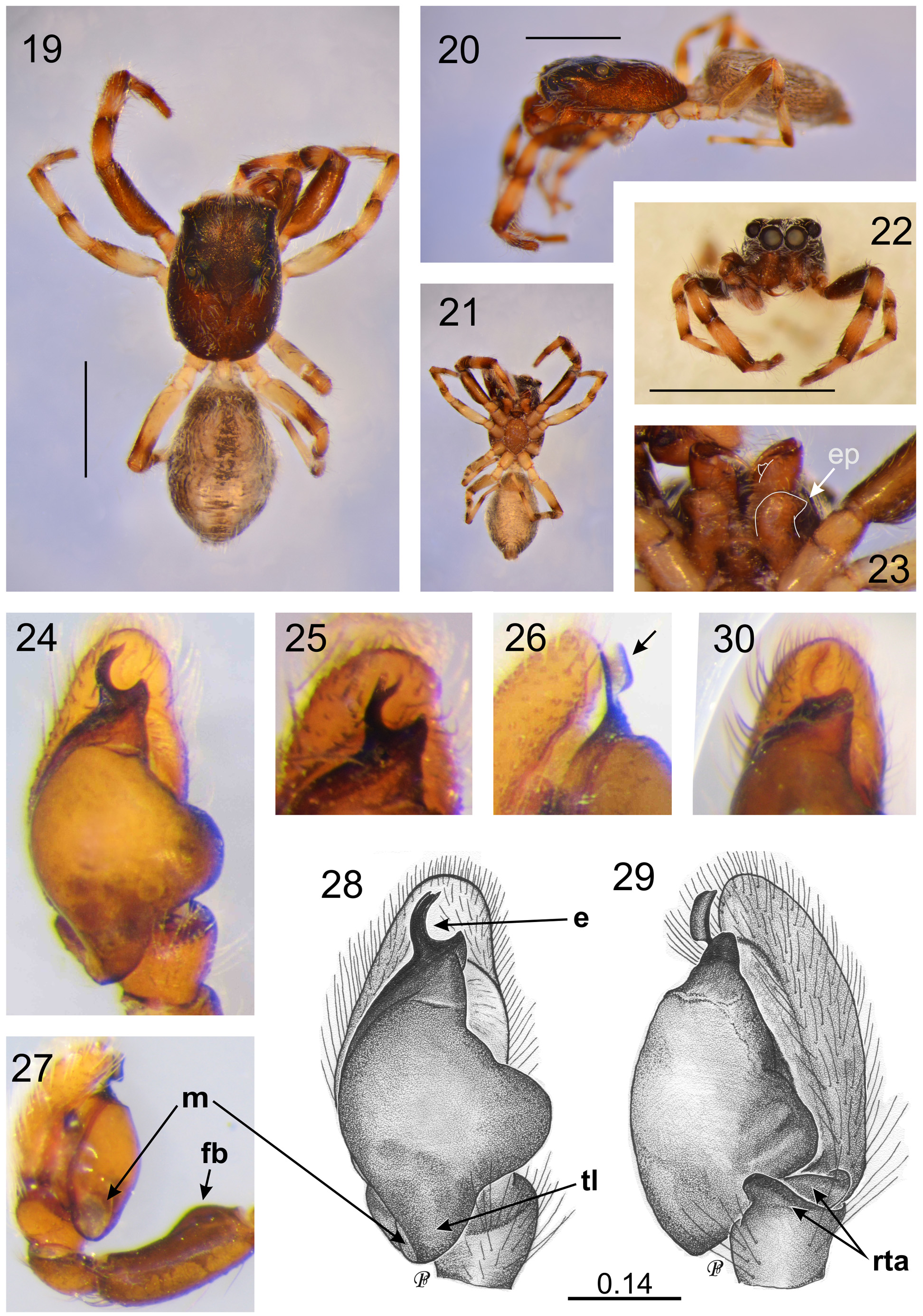
Diagnosis. The genus is distinguished by the following combination of characters: male palpal organ with a femoral bump ( Figs 7 View FIGURES 1 – 10 , 27 View FIGURES 19 – 30 , 46 View FIGURES 39 – 48 ), similar to that of a couple of unrelated Australian genera, namely Grayenulla Żabka, 1992 and Pseudosynagelides Żabka, 1991 , and in Heliophanus C.L. Koch, 1833 , genus not present in Australia ( Wesołowska 1986; Żabka 1991, 1992). Posterior lobe of bulbus with a unique membranous structure ( Figs 7 View FIGURES 1 – 10 , 27 View FIGURES 19 – 30 , 46 View FIGURES 39 – 48 ). Embolus arises as the continuation of bulbus and is sometimes accompanied by membranous pars pendula ( Fig. 8 View FIGURES 1 – 10 ), like in possibly related Menemerus Simon, 1868 (see e.g. Davies & Żabka 1989, fig. 55, marked as membranous conductor). Epigyne with a large caudal lobe/scape ( Figs 17 View FIGURES 11 – 18 , 37 View FIGURES 31 – 38 ), also present in single species of unrelated Australian Astieae and Dioleniae (see Davies & Żabka 1989, figs 19, 21) and in some species of Phintella , genus not recorded in Australia (e.g. Żabka 1985). Internal female genitalia (vulva) similar to that of Menemerus , with two (proximal and distal) chambers connected by a straight duct ( Figs 18 View FIGURES 11 – 18 , 38 View FIGURES 31 – 38 ). Spermatheca coiled in distal chamber ( Figs 18 View FIGURES 11 – 18 , 38 View FIGURES 31 – 38 ).
Description. Small, slender and flattened fissident spiders ( Fig. 1 View FIGURES 1 – 10 ), 2.8–3.5 mm long. Thoracic part much longer than cephalic (EFL <42% of CL); fovea located well behind the eye field, in the middle of thorax. Abdomen ovoid-elongate, dark, with lighter markings or chevrons. Spinnerets not distinctive. Clypeus very narrow (<7% of AME diameter); chelicerae vertical or slightly projecting, with two single prolateral teeth and one fissident retrolateral tooth. Maxillae/endites in males with protuberance ( Figs 3 View FIGURES 1 – 10 , 23 View FIGURES 19 – 30 , 43 View FIGURES 39 – 48 ). Labium wider than long (<50% of endites’ length). Sternum and venter not distinctive. Fourth legs the longest, followed the first, more robust than others, especially in males. Legs 3 and 2 shorter (for leg formula, see species’ descriptions). First tibiae and metatarsi with pairs of strong ventral spines (2-2-2 and 2-2, respectively). Bump on palpal femur only slightly marked ( P. nasalis , P. wandae ) or in the form of an apophysis ( P. occidentalis ). Palpal tibia complicated with different shapes of apophyses (see species descriptions). In P. nasalis , there is a cymbial flange. Seminal duct not meandering. Epigyne with single ( P. nasalis ) or bilobate scape ( P. wandae ). Copulatory openings oriented laterally ( P. nasalis ) or towards each other ( P. wandae ).
Relationships. For many decades after Simon’s monumental works (1901-1903), the discussion on salticid relationships was based upon morphological characters and resulted in several important works by Wanless, Galiano, Prószyński and others (review in Maddison et al. 2014). Within the last decade or so, molecular methods have widely been applied, mostly by Prof. Wayne Maddison and his team ( Maddison et al. 2008; Zhang & Maddison 2013; Maddison et al. 2014) and have brought unexpected results. The multi-gene analyses of nonsalticoid clades (cocalodines, lapsines, Spartaeinae ) basically supported some earlier morphology-based views, but the results for salticoids appeared revolutionary ( Maddison et al. 2014), proving that the higher salticid taxonomy has to be verified using molecular methods. For now, there is only morphological data available for the genus Proszynellus , and following Maddison et al. (2008), it can be provisionally placed within the Heliophaninae clade, with the genus Menemerus Simon being especially close. Both share similar palpal tibiae, more or less marked cymbial flange and membranous pars pendula. Also, in some Menemerus species the internal female genitalia have strongly sclerotised chambers with thin coiled ducts inside ( Wesołowska 1999). Three other genera, Heliophanus , Phintella Strand, 1906 and Pseudicius Simon, 1885 ( Andreeva et al. 1984; Wesołowska 1986), seem related, too. In the first, femoral apophysis, bulbus and embolus are similar. The scape/caudal lobe, so distinctive in Proszynellus , can also be found in many species of Phintella (see Żabka 1985), but at this stage it is hard to judge potential homologies.
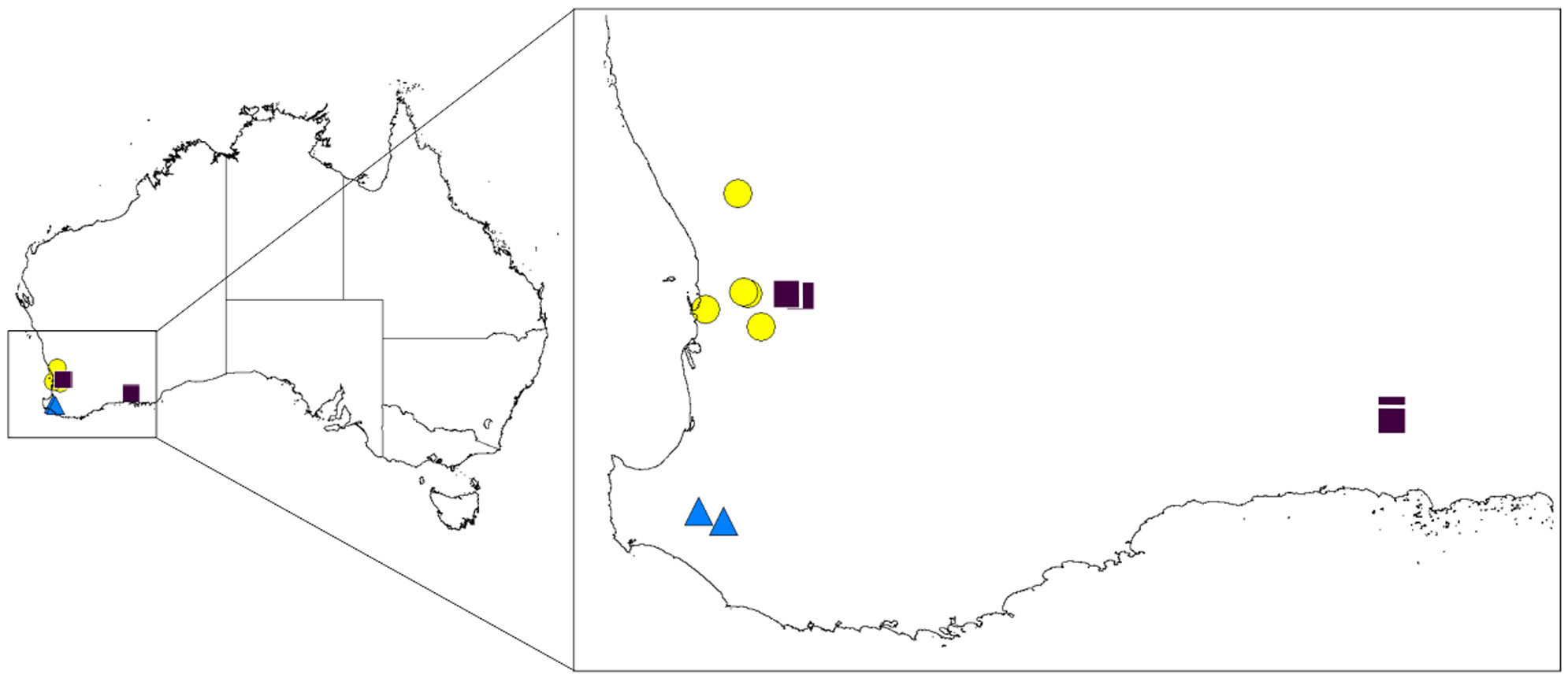
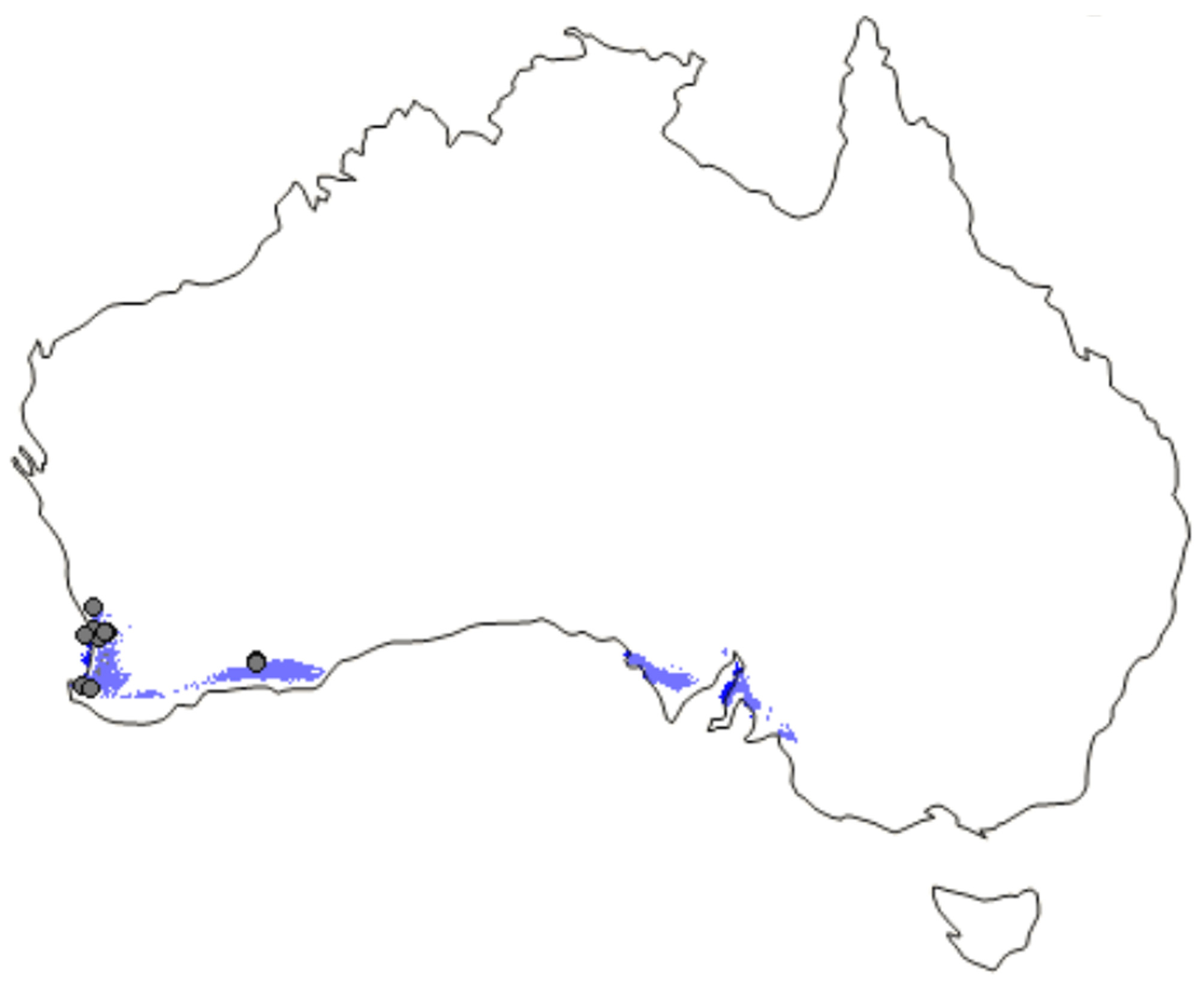
Distribution. The distributional records of Proszynellus are (so far) limited to localities in Western Australia ( Fig. 49 View FIGURE 49 ), although the predicted distribution suggests a wider range, including South Australia ( Fig. 50 View FIGURE 50 ).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.