
MYRIOPATHIDAE OPRESKO, 2001B
|
publication ID |
https://doi.org/10.1111/zoj.12060 |
|
persistent identifier |
https://treatment.plazi.org/id/DE793A5A-FFBA-ED41-1113-FEED85B6FA33 |
|
treatment provided by |
Marcus |
|
scientific name |
MYRIOPATHIDAE OPRESKO, 2001B |
| status |
|
MYRIOPATHIDAE OPRESKO, 2001B View in CoL
Opresko (2001b) established the family Myriopathidae for a group of species previously classified in the Antipathidae . Compared with antipathids, myriopathids have smaller polyps, shorter tentacles with rounded tips, and forked or antler-like spines on the base of the stem and holdfast. The Myriopathidae display a moderate amount of morphological variation in the growth form of the corallum. Colonies may be irregularly branched or pinnulate, and take on bottlebrush, irregularly branched or flabellate shapes; there are no unbranched forms known. The polyps tend to be slightly elongated in an axial direction and generally not more than 1 mm in transverse diameter. The spines are typically conical to bladelike on the smallest branches and pinnules, becoming longer and more acicular (needle-like) on the larger branches and stem. Spines increase in density on larger branches and stem; often bifid, trifid or dendritic. Spines can be smooth-surfaced or ornamented with very fine papillae. The family comprises 34 species in five genera.
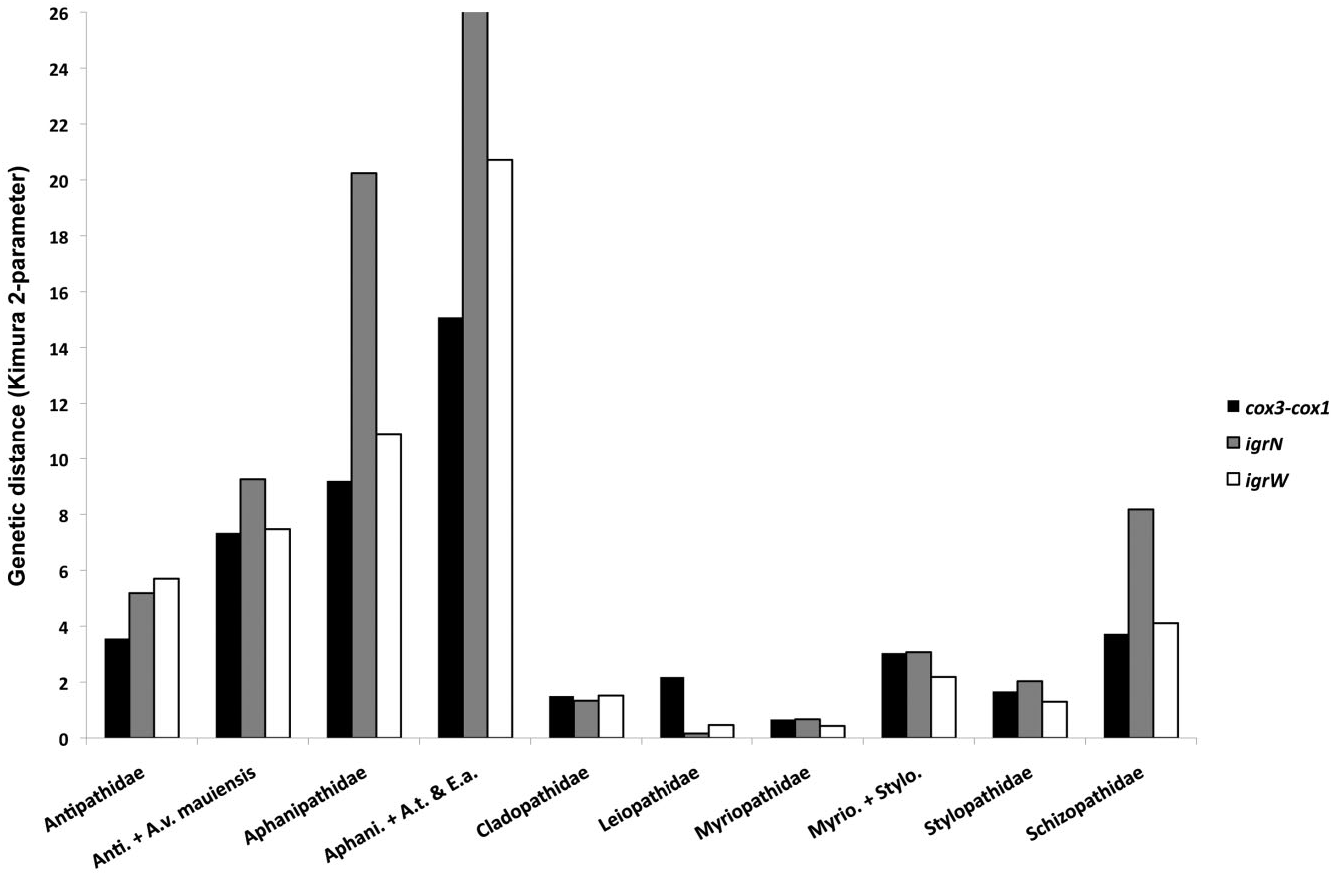
Based on morphological analyses, five species, representing three genera, were obtained for sequencing, of which four could be referred with certainty to valid nominal species ( Table 1 & Supporting information Table S1). In all phylogenetic analyses, the myriopathids clustered into a monophyletic clade, with the exception of the PhyloBayes analysis of the mt-contig based on the full alignment (not shown), where these species formed a polytomy with stylopathid species. The close relationships among – and the low genetic distances between – stylopathids and myriopathids are sufficient to warrant consideration for combining them into a single family (see discussion below). Pairwise comparisons of mt sequences between Myriopathidae species revealed genetic distances lower than seen in other families ( Fig. 1 View Figure 1 ; maximum distance for cox 3- cox 1: 0.66%, igrN: 0.67%, igrW: 0.43%). Not surprisingly, the highest genetic distances were for comparisons between the most geographically distant taxa: Myriopathes ulex ( Ellis & Solander, 1786) from Hawaii and Gulf of Mexico species in the genera Plumapathes Opresko, 2001b and Tanacetipathes . There was very little divergence among the Atlantic myriopathids, even between genera. We sequenced two colonies each from Plumapathes pennacea [both from the Flower Garden Banks (FGB) in the Gulf of Mexico], Tanacetipathes barbadensis Brook, 1889 (both from FGB), and T. tanacetum ( de Pourtalès, 1880) (one from FGB, the other from the Atlantic coast of northern Florida, USA). None of the colonies differed at igrW, but four igrN / cox 3- cox 1 combined haplotypes were observed. Unexpectedly, these haplotypes differentiated the two colonies within each species, i.e. intra specific variation, but were shared among the species and genera (Supporting information Table S1). For example, the colony of T. tanacetum from northern Florida had the same igrN / cox 3- cox 1 combined haplotype as colonies of T. barbadensis and Plumapathes pennacea , both from FGB. A seventh colony, from the Gulf of Mexico off Louisiana and identified as Tanacetipathes cf. paula Pérez & Costa, 2005 , shared an igrN haplotype with an FGB P. pennacea (USNM 1086302) ( T. paula is considered by Loiola & Castro, 2005 to be a synonym of T. thamnea but considered by the present authors to be morphologically distinct). Again, these data are an example of the extremely low rates of substitution in the mitochondrial genes of black corals, perhaps combined with relatively recent divergence of the Atlantic species.
The S + M ( Stylopathidae + Myriopathidae ) complex grouped sister to the aphanipathid Elatopathes abietina (USNM 1116469), although with conflicting support from the different tree-building algorithms (BS: 56.7–59.4, BPP: 62–100; placement of the S + M complex in the PhyloBayes analysis of the Gblocksedited mt-contig alignment was unresolved). Again, Bayesian analysis of the full (BPP: 93) and GBlocksedited (BPP: 95) alignment of igrN alone resulted in strong support for E. abietina grouping sister to the S + M complex. All Myriopathidae colonies examined contained a group I intron within cox 1 (see discussion below).
Although including only a single representative of the Myriopathidae [ T. barbadensis (USNM 1116465)] and a single representative of the Stylopathidae [ Stylopathes sp. (NIWA 16047)], the nuc-contig phylogenies recovered, with strong support, the relationship of the S + M complex (BS and BPP: 100), which grouped sister to E. abietina (BS: 90.6; BPP: 68–99).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.