
Pheidole simplispinosa Sarnat
|
publication ID |
https://doi.org/10.5281/zenodo.182030 |
|
DOI |
https://doi.org/10.5281/zenodo.6234379 |
|
persistent identifier |
https://treatment.plazi.org/id/DF17786E-FF8E-FFE4-53C3-2ACBFC2BDC20 |
|
treatment provided by |
Plazi |
|
scientific name |
Pheidole simplispinosa Sarnat |
| status |
sp. nov. |
Pheidole simplispinosa Sarnat sp. n.
Figs. 47–49 View FIGURES 32 – 52 , 68–70 View FIGURES 53 – 73 , 89–91 View FIGURES 74 – 93
Holotype major, FIJI: Koro I., Mt. Kuitarua 3.7 km NW Nasau Village, 20.vi. 2005, 470m, -17.29083°, 179.40183°, primary rainforest, nesting in soil, (E. M. Sarnat), EMS#2084, CASENT0171106 ( FNIC).
Paratypes. From same nest series as holotype: 1 dealate queen (CASENT0171107), 4 majors (CASENT0174208, CASENT0174209, CASENT0174211, CASENT0174212), 3 minors (CASENT0171022, CASENT0174210, CASENT0174213), ( FNIC, NMNH, ANIC); additional specimens in alcohol ( NMNH).
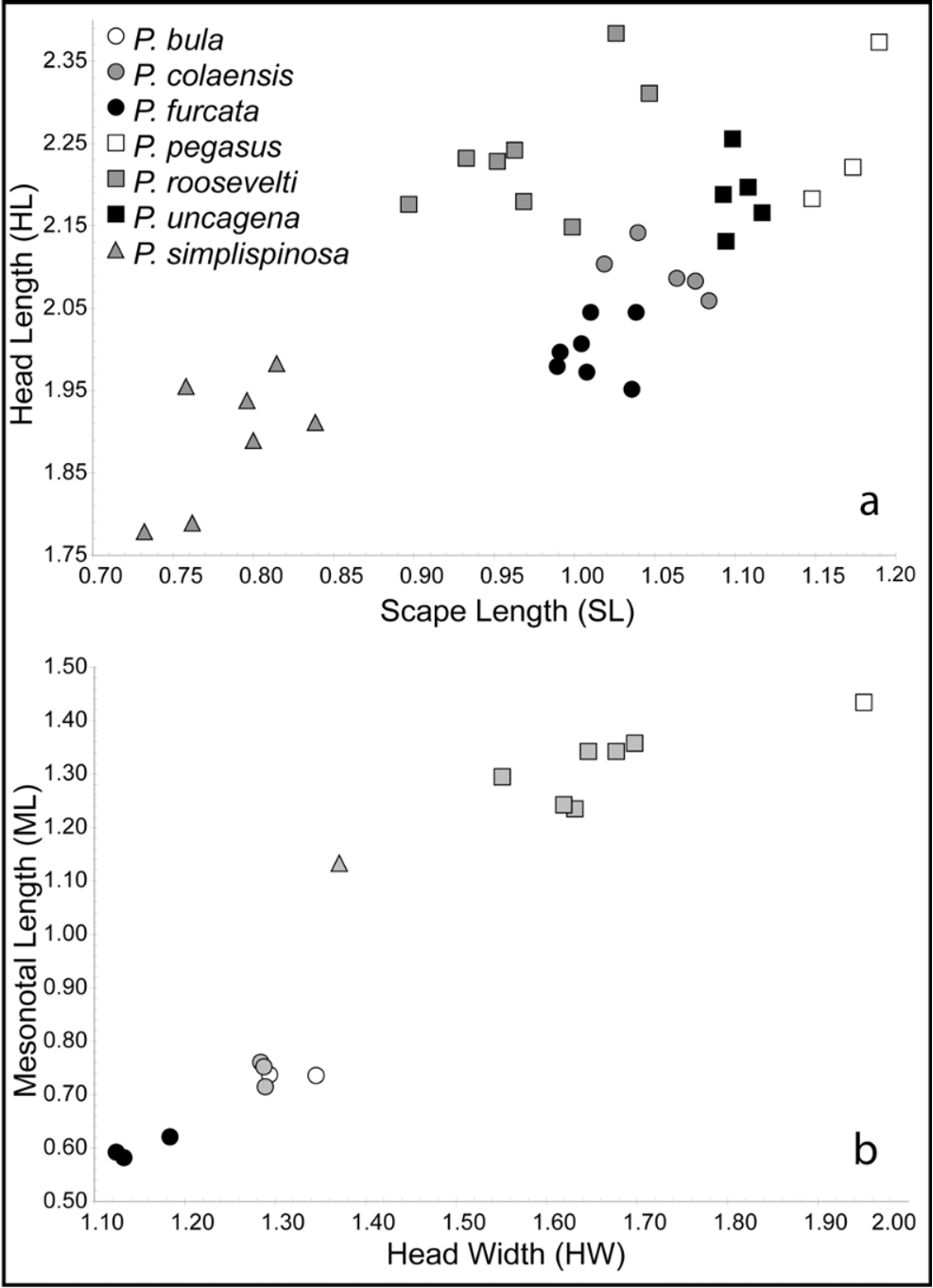
MAJOR. TL 5.65–6.78, HL 1.78–1.96, HW 1.64–1.84, CI 0.89–0.93, FL 1.09–1.22, FI 0.60–0.64, SL 0.73–0.84, SI 0.39–0.44 (7 measured).
Head with sides not distinctly broader posterior to eyes than anterior to eyes. Median ocellus occasionally present. Scapes short. Mesonotal process, in profile, truncated into a blunt process without lamellate posterior margin; in dorsal view posterior margin flat to convex. Propodeal spines simple, evenly tapering to a single straight acuminate point without becoming bifurcate or angulate apically. In posterior view, petiole node dorsum strongly excised, sides moderately excised. Postpetiole taller than long, as tall as petiole, steep anterior and posterior faces converging to form an obtusely angulate vertex; in dorsal view subpentagonal with strong lateral projections.
Region between frontal carinae with parallel longitudinal carinae extending posteriorly and arcing onto posterolateral corners. Posterolateral lobes, in full face view, with distinct transverse rugae extending from median cleft to posterolateral corners. Sculpture of vertex and posterolateral lobes varies from weakly to densely rugoreticulate. Intercarinular spaces vary from densely foveolate to smooth and shining. Rugoreticulum present between eye and antennal insertion. Antennal scrobe mostly smooth and shining to strongly foveolate. Clypeus smooth and shining; anterior margin without carinae. Lateral and ventrolateral portions of posterolateral lobes lightly sculptured to entirely smooth and shining; posteriorly with transverse striae, discontinuous rugae and shallow oblong impressions. Head venter rugose. Pronotal sculpture varies from dense foveolate ground sculpture overlain by transverse striae to weak foveolate ground sculpture overlain by weak transverse striae. Mesonotum, in dorsal view, varies from foveolate and striate to smooth and shining. Anepisternum weakly rugose. Katepisternum weakly striate. Petiole with apical and posterior faces smooth, laterally and ventrally rugose. Postpetiole dorsum mostly smooth and shining, sides rugose. Basigastral costulae weakly to moderate with interspaces smooth and shining to foveolate; sternite of first gaster segment lightly sculptured laterally. Gaster otherwise smooth and shining. Body reddish-brown with lighter appendages.
MINOR. TL 2.96–3.41, HL 0.66–0.75, HW 0.65–0.74, CI 0.98–1.03, FL 0.80–0.91, FI 1.18–1.29, SL 0.77–0.85, SI 1.10–1.23, AE 0.25–0.32, DE 0.00, PSI 0.00 (10 measured).
Head, in full face view, subcordate, sides convex, posterolateral corners rounded and obtuse, posterior margin concave; in profile, posterolateral corners not pinched. Genal carinae very short and weakly elevated on ventrolateral portion of head. Clypeus with anterior margin convex laterally, concave medially. Frontal carinae terminating before eye level. Mesonotal process truncated into a short process without lamellate or distinct margin; in dorsal view, attachment to mesonotum broad, posterior margin evenly convex. Propodeal spines simple, taper evenly to an acuminate point without distal angle or bifurcation.
Head mostly smooth and shining to latitudinally rugulose with densely packed foveolate ground sculpture; ventrally smooth and shining. Clypeus with a few weak carinae attached to anterolateral border. Promesonotum, in dorsal view, transversely rugulose with lightly impressed to densely packed foveolate ground sculpture. Mesopleuron mostly smooth and shining to foveolate. Reddish-brown with paler appendages.
QUEEN. TL 6.82, HL 1.29, HW 1.37, CI 1.06, SL 0.87, SI 0.68, FL 1.18, FI 0.91, ML 0.83, MI 1.05 (1 measured).
Head subquadrate with sides approximately as broad anteriorly as posteriorly. Mesoscutum, in profile, large, greater than half the height of pronotum; in dorsal view, mostly obscuring pronotum. Scutellum, in dorsal view, with posterior portion produced as a weakly elevated subtriangular plate. Propodeal spines simple, evenly tapering to a single straight acuminate point without becoming bifurcate or angulate apically. Petiole broadly cuneate; in posterior view, petiole node dorsum weakly concave. Postpetiole, in dorsal view, subpentagonal with moderate lateral projections.
Region between frontal carinae with crenulated longitudinal and occasionally branching carinae reach posterior margin. Region between eyes and antennal insertions with elevated crenulated carinae that reach posterolateral corners; intercarinular spaces densely packed with overlapping foveolae. Head venter rugoreticulate. Antennal scrobe foveolate. Clypeus mostly smooth and shining; anterior margin with short carinae laterally, median carinae present. Pronotum rugoreticulate. Mesoscutum, in dorsal view, with straight parallel carinae that run posteriorly towards median. Scutellum mostly smooth and shining. Anepisternum finely rugoreticulate. Katepisternum weakly striate. Petiole rugose. Postpetiole rugulose. First tergite of gaster longitudinally sculptured basally, sculpture immediately posterior to postpetiole longer than length of postpetiole. First sternite of gaster sculptured basally. Reddish-brown with lighter appendages.
Etymology. The specific epithet simplispinosa is a noun in apposition derived from the combination of the Latin simplex, meaning simple, and spina, meaning spine.
Diagnosis, distribution and biology. Pheidole simplispinosa is the most distinctive of all P. roosevelti - group species. It is the only member of the group in which the spines are simple and evenly straight without becoming modified into distal angles or bifurcations. The mesonotal process so prominent in other all other P. roosevelti -group species is truncated into a blunt process such that the angle between the dorsal face of the mesonotum and the mesonotal declivity is obtuse (i.e.,>90°). Beyond the simplified spine and mesonotal process, P. simplispinosa is also the smallest of this group and has the shortest limbs relative to its size. The queens of P. simplispinosa , like those of P. roosevelti and P. p e g a s u s, are characterized by their well-developed mesosomas.
Although single turret nests were observed, the species is also capable of constructing nests with multiple entrances. One such nest, from Mt. Delaikoro on Vanua Levu , was composed of irregular mounds of excavated soil.
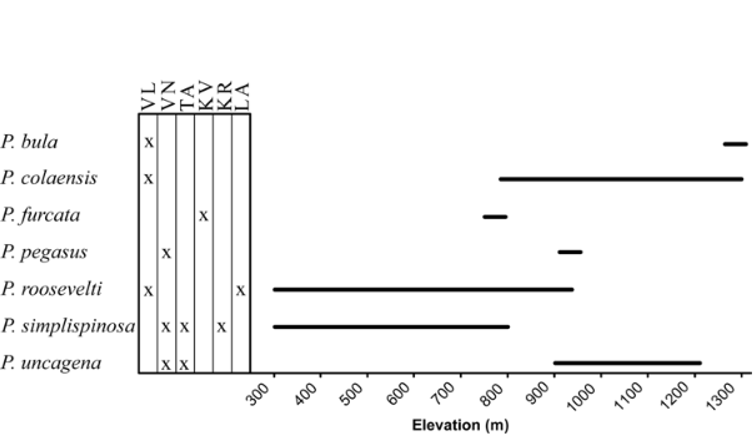
Pheidole simplispinosa has a range within the Fiji archipelago rivaled only by P. roosevelti . Like P. roosevelti , this species tolerance of lower elevation habitat ( Fig. 95 View FIGURE 95 ), may serve as some explanation for its wide range. The two species, however, are entirely allopatric. Whereas P. roosevelti claims the more southern islands of Viti Levu and Ovalau, P. simplispinosa occurs in the northern islands of Vanua Levu, Taveuni and Koro.
A significant variation in sculpture is associated with the geography of P. simplispinosa . The Koro material, including the type series, exhibits the strongest rugulae on the face and body and the most dense, well defined foveolate ground sculpture. The Taveuni specimens, with only faint hints of facial foveolae and weak mesosomal rugulae, occupy the opposing end of the phenotypic spectrum. Were it not for the intermediate Vanua Levu material, these contrasting morphologies might argue for the designation of separate species. Fortunately, enough collections of P. simplispinosa have been made on Vanua Levu to study morphological variation on the longitudinal axis. When specimens are arranged according to longitude, it becomes apparent that the strength of sculpture diminishes eastwards as the collections approach Taveuni. If sculpture strength can be used as a surrogate for relationship, it suggests that more gene flow occurs between populations from western Vanua Levu and Koro , and eastern Vanua Levu and Taveuni , than between Koro and Taveuni.
While further study of the population structure of P. simplispinosa may reveal illuminating patterns concerning phylogeographic patterns within the archipelago, perplexing questions surround the evolution of this species within the larger context of the P. roosevelti -group. Pheidole simplispinosa , with its simple propodeal spines, truncated mesonotal process, and divergent anatomical proportions ( Fig. 94 View FIGURE 94 ) does not immediately suggest itself as a close relative to any of the other P. roosevelti -group species.
Several hypotheses can be proposed to explain the relationship of P. simplispinosa to the other members of the group. The first hypothesis submits that P. s i m p l i s p i n o s a is sister to all other members of the P.
roosevelti -group. If this hypothesis is supported, it allows for the possibility that the P. roosevelti -group descended from a hypothetical ancestor that, with neither modified propodeal spines nor a mesonotal process, more closely resembled typical Pheidole species. The second hypothesis submits that P. simplispinosa is nested within the P. roosevelti -group. If P. simplispinosa is, in fact, nested within the group, it predicts that the species is derived from an ancestor that bore modified propodeal spines and an extended mesonotal process. In this scenario, the secondary reductions of propodeal spines and mesonotum render the similarities between P. simplispinosa and Pheidole species outside the P. roosevelti -group homoplasy.
A thorough multi-gene phylogeny with appropriate outgroup taxa (including quadrispinosa -group, quadricuspis -group and cervicornis -group species) will help to resolve both population and species level relationships of this enigmatic ant, and serve as a tool for testing the proposed hypotheses.
Additional material examined. FIJI. Koro: Mt. Kuitarua 3.1 km WNW Nasau Village, 20.vi. 2005, 440m, -17.29528°, 179.40433°, primary rainforest, nesting in dead tree fern, (E. M. Sarnat), EMS#2097, 3 majors (CASENT0174223 – CASENT0174225); Mt. Kuitarua 4 km WNW Nasau Village, 7.iii. 2003, 380m, - 17.29528°, 179.40433°, lowland rainforest, from sifted leaf litter, (E. M. Sarnat), FJKR 312_K01, 9 minors (CASENT0174226 – CASENT0174234); Mt. Nabukala 5.0 km WSW Nasau Village, 15.iii. 2005, 520m, - 17.31250°, 179.38617°, primary rainforest, in soil, (E. M. Sarnat), EMS#1906, 6 majors (CASENT0174214, CASENT0174215, CASENT0174217, CASENT0174218, CASENT0174220, CASENT0174221), 3 minors (CASENT0174216, CASENT0174219, CASENT0174222); Vanua Levu : 2 km NNW Kasavu Village, 28.viii. 2003, 300m, -16.62000°, 179.83333°, lowland rainforest, from sifted leaf litter, (A. Rakabula), FJVN 330_K01, 1 minor (CASENT0174251); 2 km NNW Kasavu Village, 29.viii. 2003, 300m, -16.71639°, 179.66333°, lowland rainforest, from sifted leaf litter, (A. Rakabula), FJVN 330_K02, 1 minor (CASENT0174252); Mt. Delaikoro 3.7 km SE Dogoru Village, 31.viii. 2006, 699m, -16.57525°, 179.31638°, mid-elevation rainforest, from sifted leaf litter, (E. P. Economo), EPE#62, 1 minor (CASENT0174253); Mt. Delaikoro 3.7 km SE Dogoru Village, 31.viii. 2006, 699m, -16.57525°, 179.31638°, mid-elevation rainforest, nesting in soil, (E. M. Sarnat), EMS#2375, 7 majors (CASENT0174240, CASENT0174241, CASENT0174243, CASENT0174244, CASENT0174246, CASENT0174248, CASENT0174249), 4 minors (CASENT0174242, CASENT0174245, CASENT0174247, CASENT0174250); Mt. Vatudiri 3km NW Waisali Village, 2.ix. 2006, 570m, -16.62905°, 179.21103°, mid-elevation rainforest, from turret nest in bare soil, (E. M. Sarnat), EMS#2396, 3 majors (CASENT0174235, CASENT0174236, CASENT0174238), 2 minors (CASENT0174237, CASENT0174239); Mt. Vatudiri 3km NW Waisali Village, 2.ix. 2006, 570m, - 16.62905°, 179.21103°, mid-elevation rainforest, from sifted leaf litter, (E. P. Economo), EPE#79, 1 minor (CASENT0174254); Taveuni: Devo Peak 3.6 km SE Tavuki Village, 17.vi. 2005, 734m, -16.83056°, - 179.97433°, garden/primary rainforest edge, from sifted leaf litter, (E. M. Sarnat), EMS#1949, 1 major (CASENT0174255), 2 minors (CASENT0174256, CASENT0174257); Devo Peak 3.6 km SE Tavuki Village, 17.vi. 2005, 734m, -16.83056°, -179.97433°, garden/primary rainforest edge, from sifted leaf litter, (E. M. Sarnat), EMS#2069, 3 minors (CASENT0174258 – CASENT0174260); Devo Peak 3.9 km SE Tavuki Village, 17.vi. 2005, 775m, -16.83278°, -179.97343°, primary rainforest edge, from sifted leaf litter, (E. M. Sarnat), EMS#2068, 3 minors (CASENT0174261 – CASENT0174263).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |