
Neoperla Needham, 1905
|
publication ID |
https://doi.org/10.11646/zootaxa.5316.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:BC922E16-2614-4F3D-AD82-87A845DE7E2B |
|
DOI |
https://doi.org/10.5281/zenodo.16763833 |
|
persistent identifier |
https://treatment.plazi.org/id/E12C876C-4A64-FF96-FF4F-FA1FFA0D0A5E |
|
treatment provided by |
Plazi |
|
scientific name |
Neoperla Needham, 1905 |
| status |
|
Genus Neoperla Needham, 1905 View in CoL View at ENA
Pseudoperla Banks, 1892 View in CoL (no type designated), not Pseudoperla Pictet, 1856: 37 View in CoL , pl. IV, figs. 9,10 (in Pictet & Hagen in Berendt, 1856, fossil Phasmatodea).
Neoperla Needham, 1905: 108 View in CoL ; replacement name, type species (by original designation): Perla occipitalis Pictet, 1841 View in CoL (redescriptions: Zwick 1987, Stark 1990).
Ochthopetina Enderlein, 1909b: 324 View in CoL ; syn. fide Hynes (1952). Type species (by original designation): N. aeripennis (Enderlein, 1909) View in CoL (redescription: Zwick 1973a: 496).
Javanita Klapálek, 1909: 224 View in CoL ; type species (designated by Klapálek, 1923b): Neoperla caligata ( Burmeister, 1839) View in CoL ( syntypes lost; a synonym of Ochthopetina View in CoL whose type species, N. aeripennis View in CoL , was originally included).
Tropidogynoplax Enderlein, 1910: 141 View in CoL ; type species (by monotypy): T. fuscipes Enderlein View in CoL = N. aeripennis (Enderlein) View in CoL , fide Klapálek 1923: 177. The type of T. fuscipes View in CoL is not available and probably lost. Enderlein emphasised similarity of habitus between T. fuscipes View in CoL and N. aeripennis View in CoL . Klapálek’s interpretation of the name seemed doubtful ( Zwick 1973a: 499) but we follow him now assuming that the particular shape of the female subgenital plate was an artifact caused by drying.
Formosina Klapálek, 1913: 117 View in CoL . No type designated. Preoccupied, not Becker, 1911.
Formosita Klapálek, 1914:118 View in CoL ; nom. nov. for Formosina Klapálek, 1913 View in CoL (not Becker, 1911); type species: Neoperla costalis ( Klapálek, 1913) View in CoL (design. Zwick 2016).
Oodeia Klapálek, 1921: 321 View in CoL ; type species (designated by Klapálek 1921): N. dolichocephala Klapálek, 1909 View in CoL (redescription as Neoperla dolichocephala View in CoL -complex: Uchida & Yamasaki, 1989).
The presence of only two ocelli was the key character of Enderlein’s (1909c) Neoperlinae View in CoL . However, Peltoperlidae View in CoL , several other Perlinae View in CoL , especially the numerous very different Neotropical Anacroneuriini ( Perlidae View in CoL : Acroneuriinae View in CoL ) also share this character. Enderlein applied the name Neoperla View in CoL only to American species and created new genera for Old World species.
A forthcoming study (P. Zwick, in prep.) will address the relations of Neoperla to the other genera of Perlinae and will revise the internal systematics of the genus which also occurs in South East Asia, Palaearctic Asia, and eastern North America.
In brief, males of Neoperla are distinguished from other Neoperlini by four primary apomorphies:
1. T7 is modified;
2. T9 has a median furrow between pilose lateral humps and bears sensilla basiconica (SB);
3. the anterior half of the hemitergites is slender and delicate as compared to the massive base and the large mediobasal callus (misinterpreted as the flattened hemitergite apex by Sivec et al. 1988); and
4. a pair of hard rounded retractor sclerites projects laterally from the basal penis opening.
In many Plecoptera , also in most Perlinae , the membranous penis assumes its specific shape only temporarily while everted by hemolymph pressure. It collapses when the pressure is released. In the tribe Neoperlini , sclerites restricting the basal penis opening prevent this and penes of Neoperlini retain their shape at rest. The infolded endophallus (often simply called the sac) with its specific spine patterns is only for copula everted by hemolymph pressure.
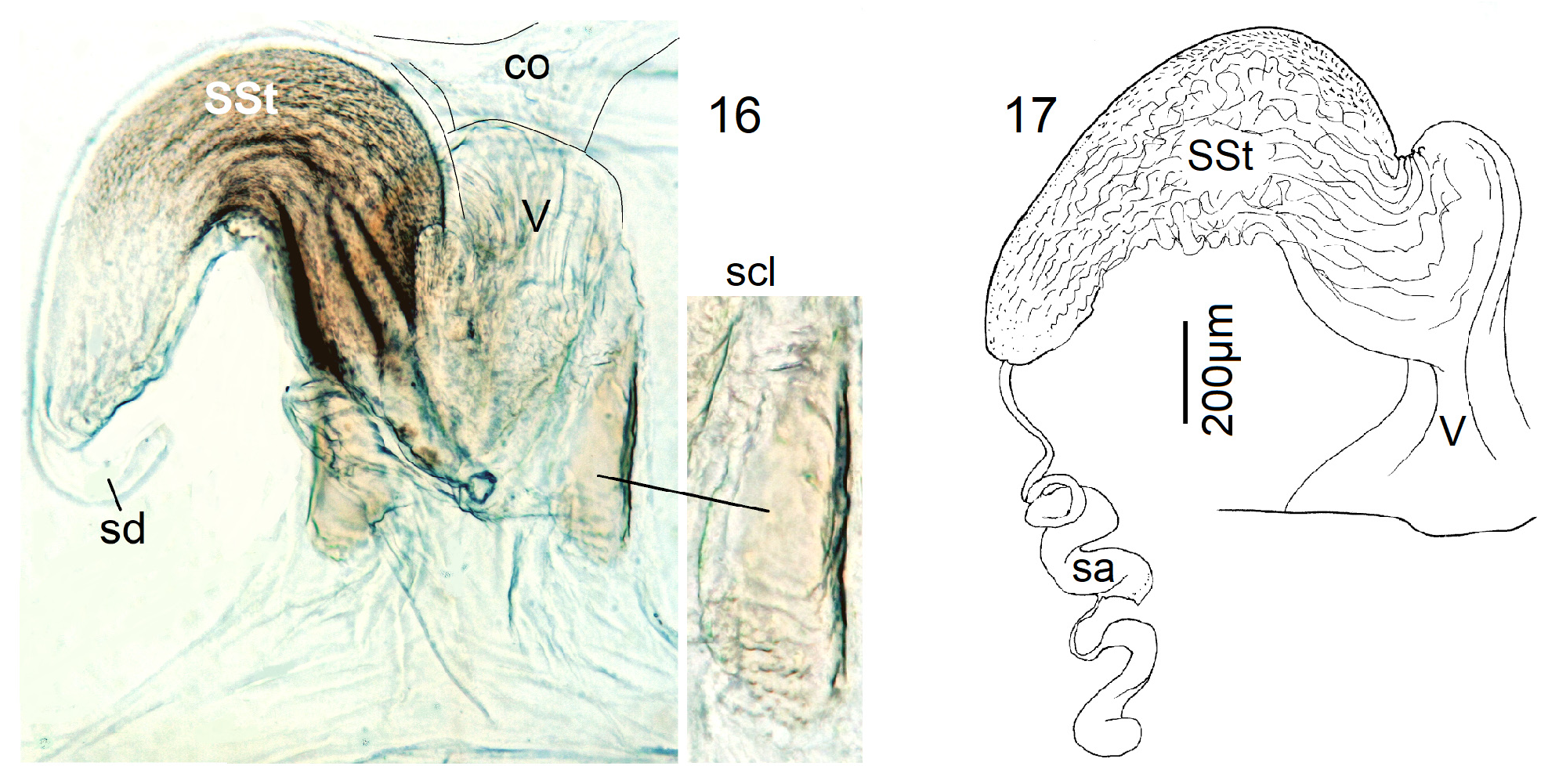
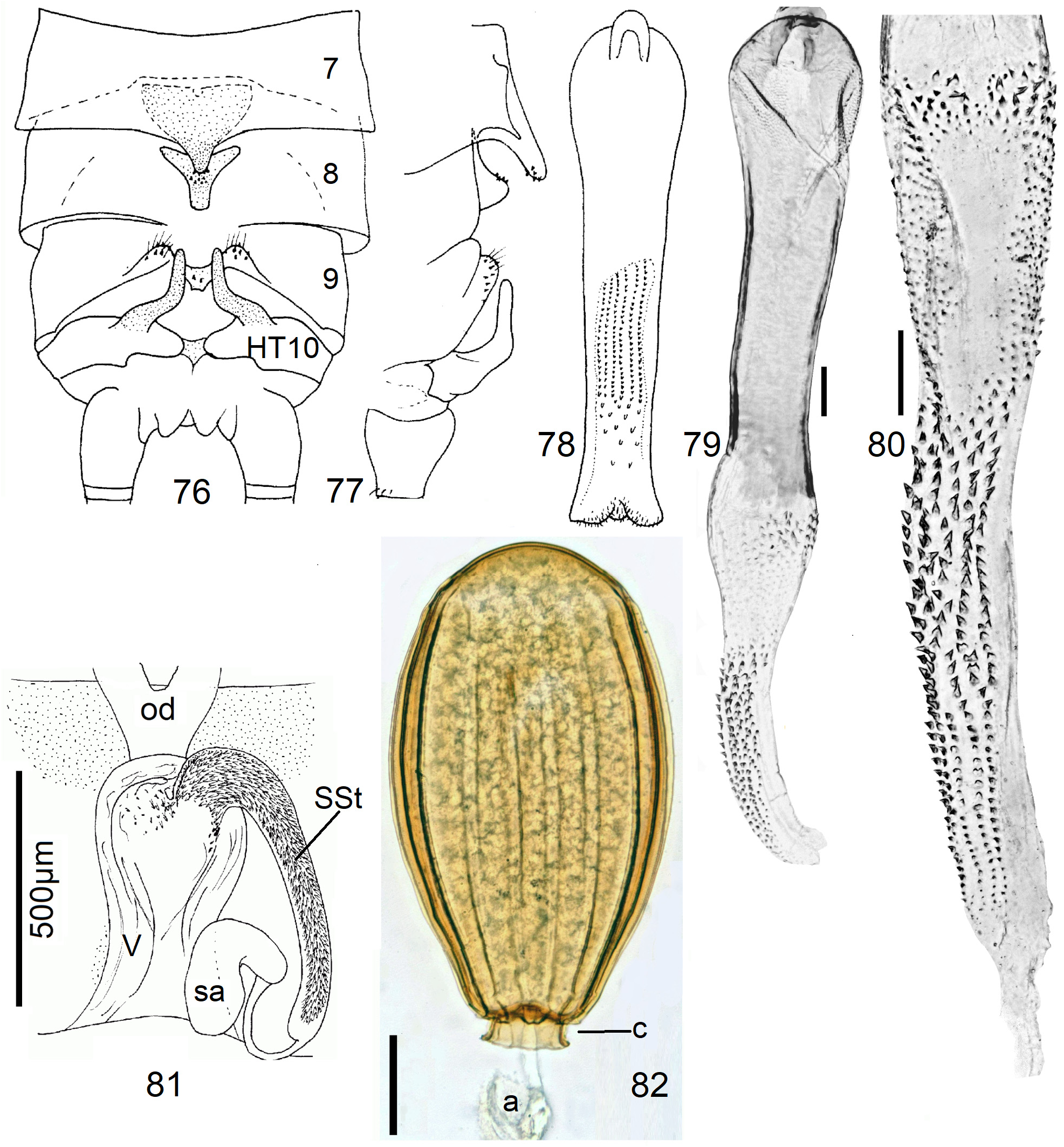
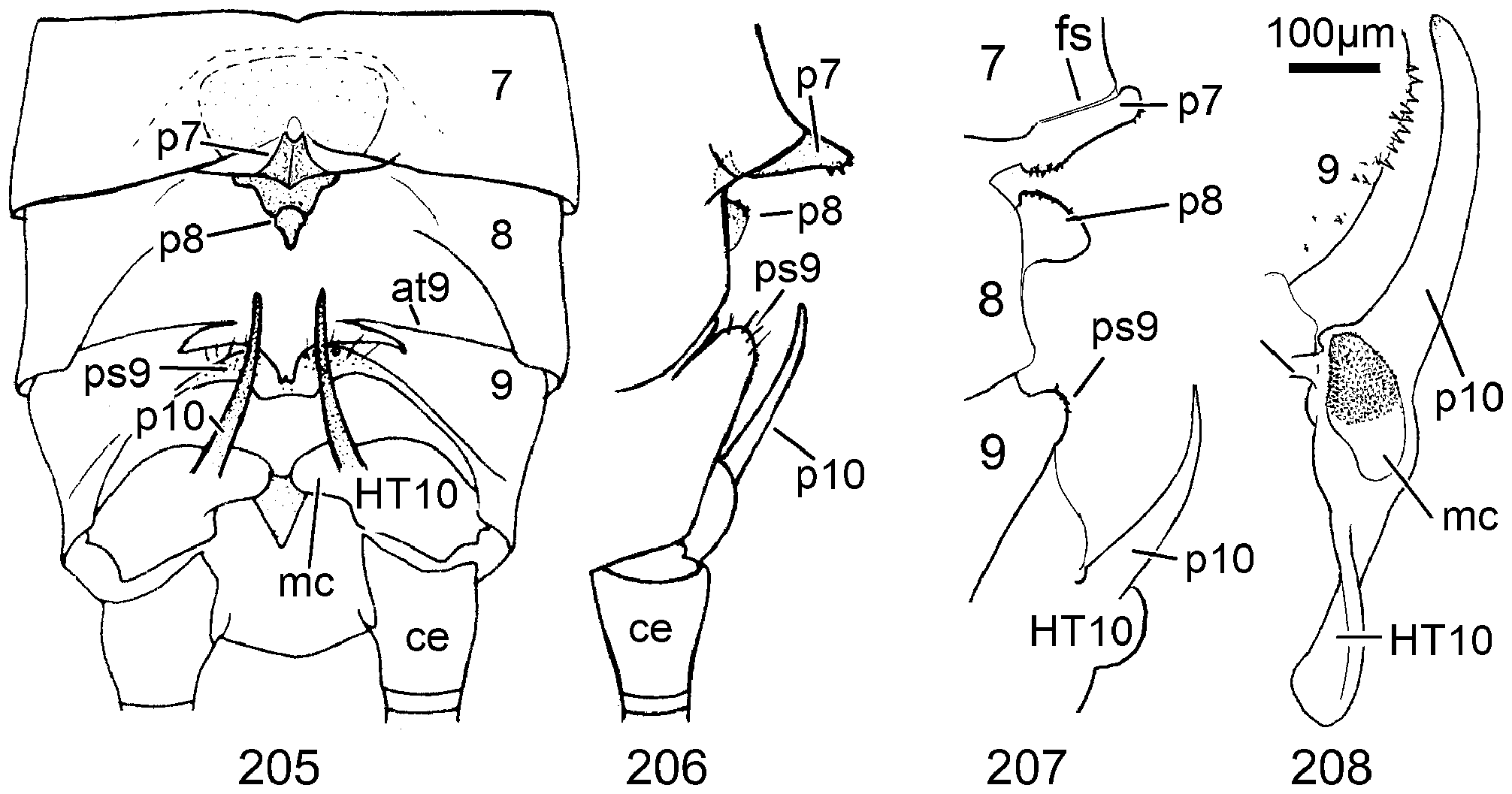
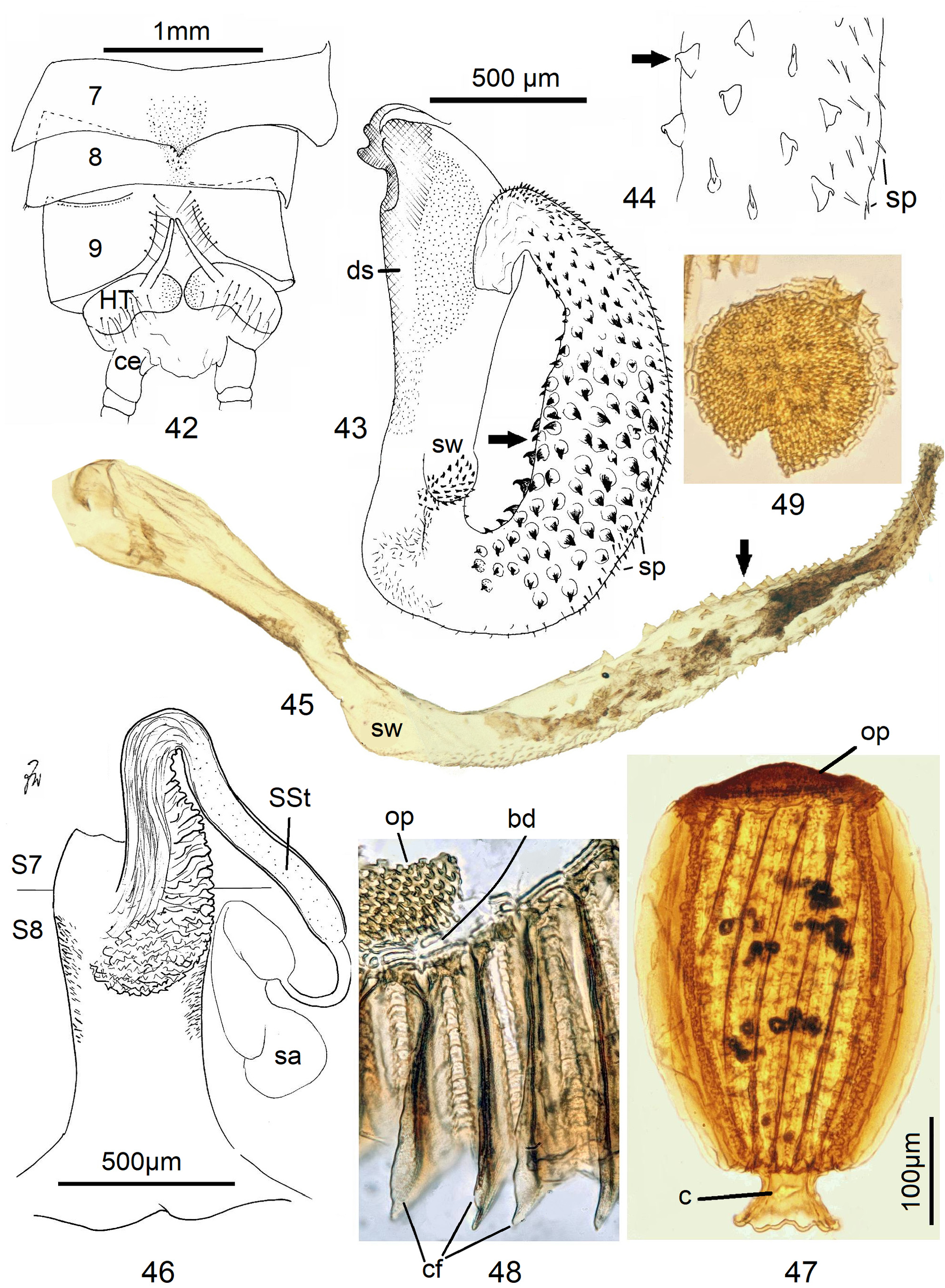
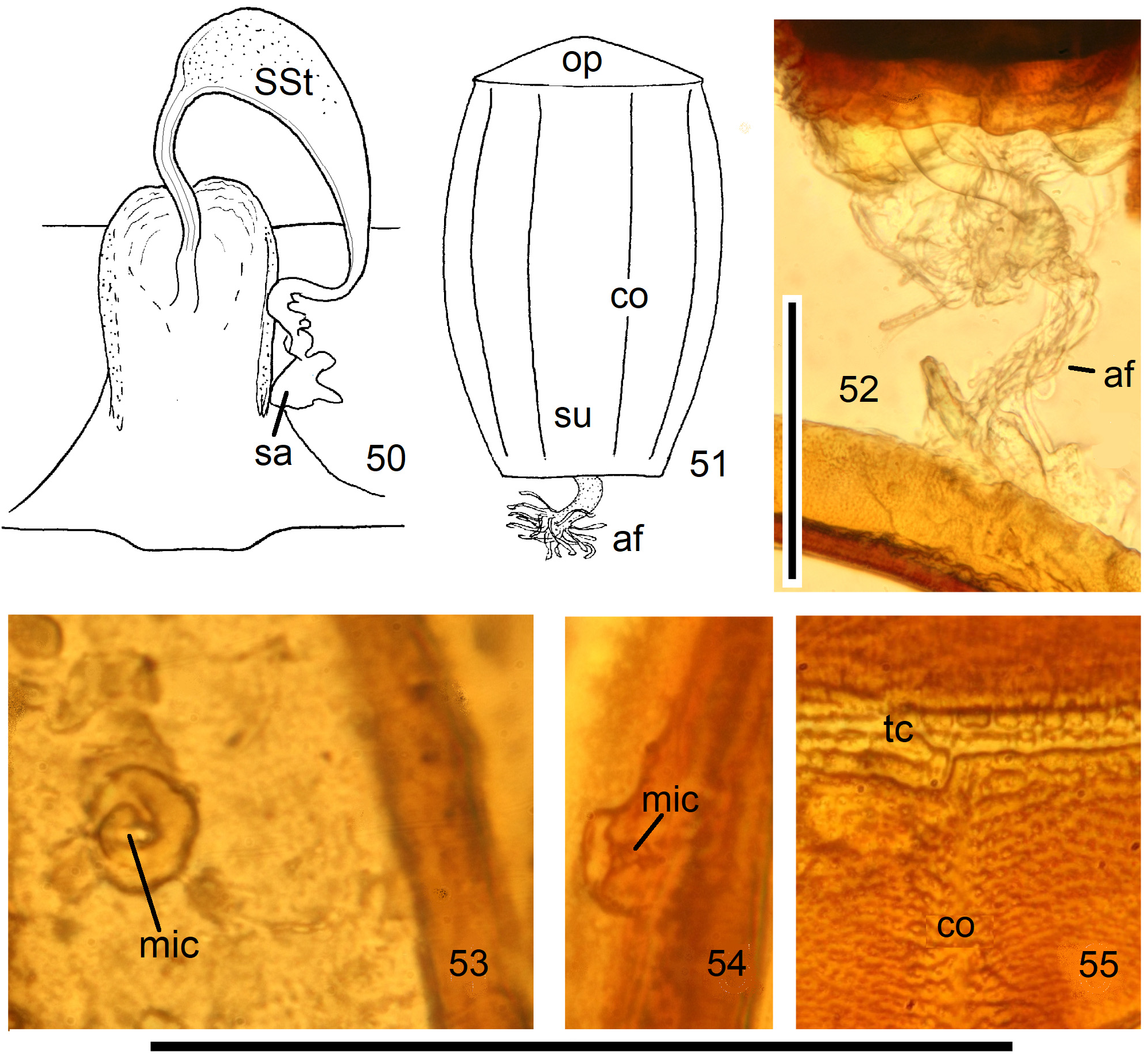
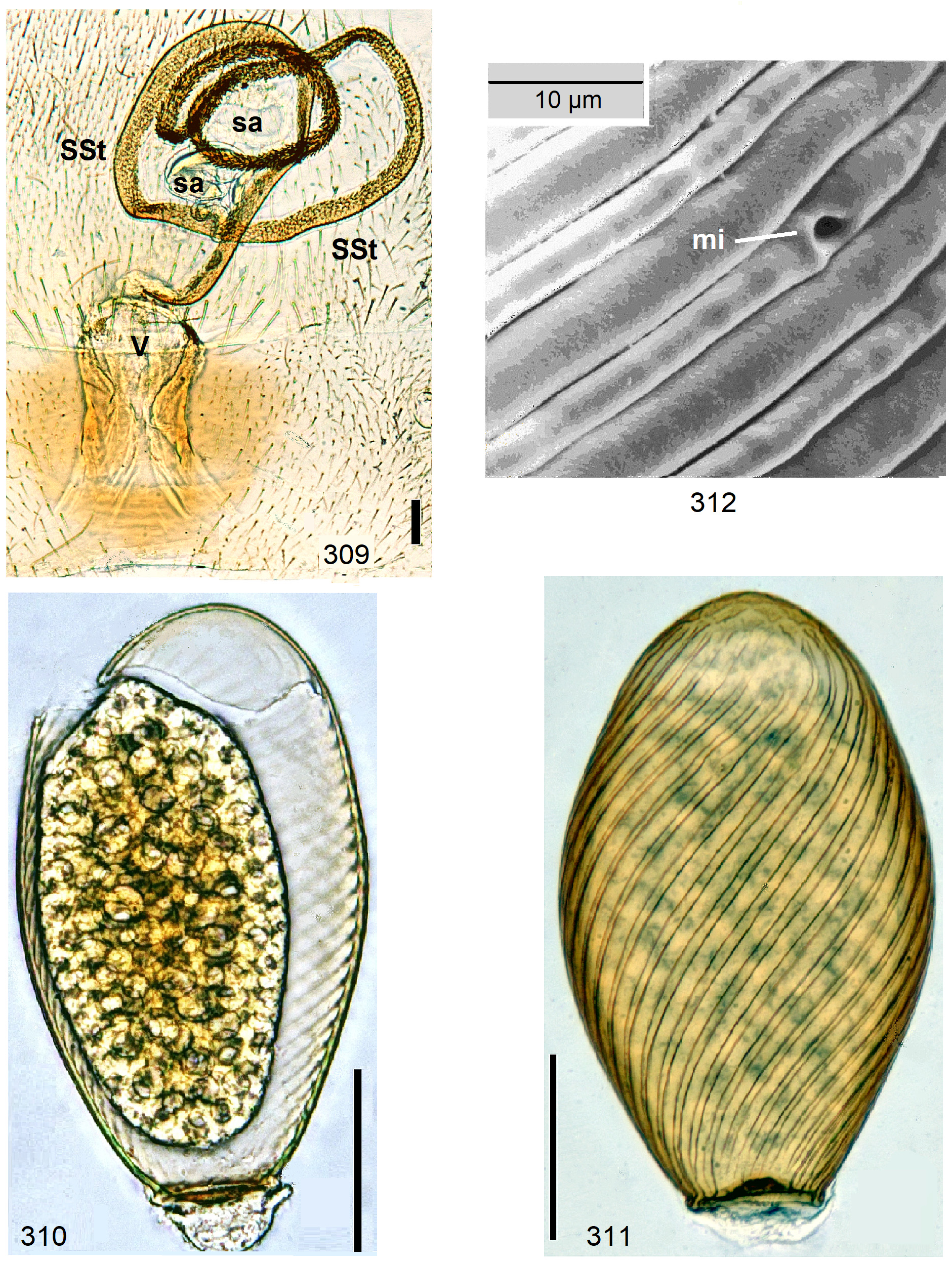
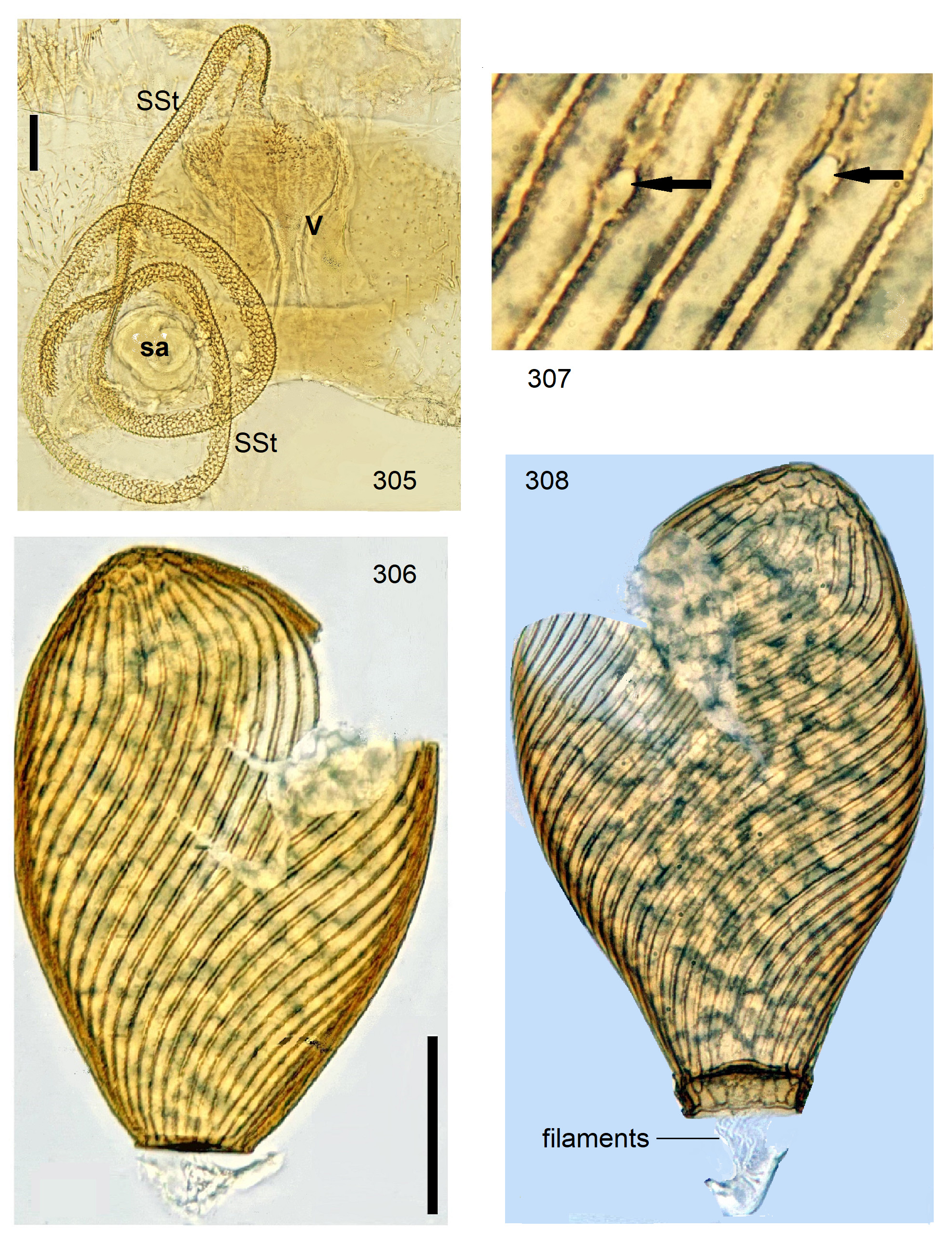
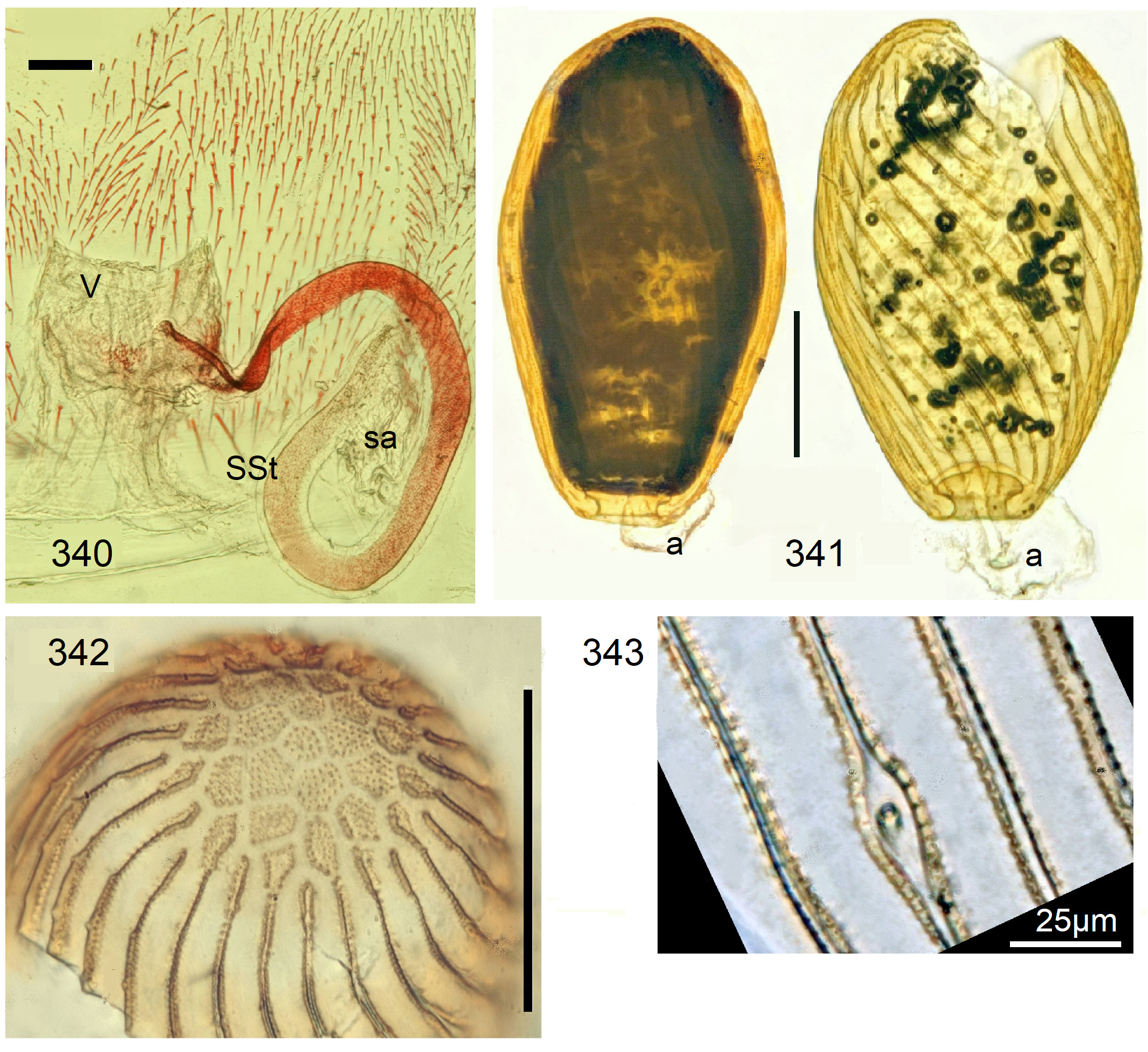
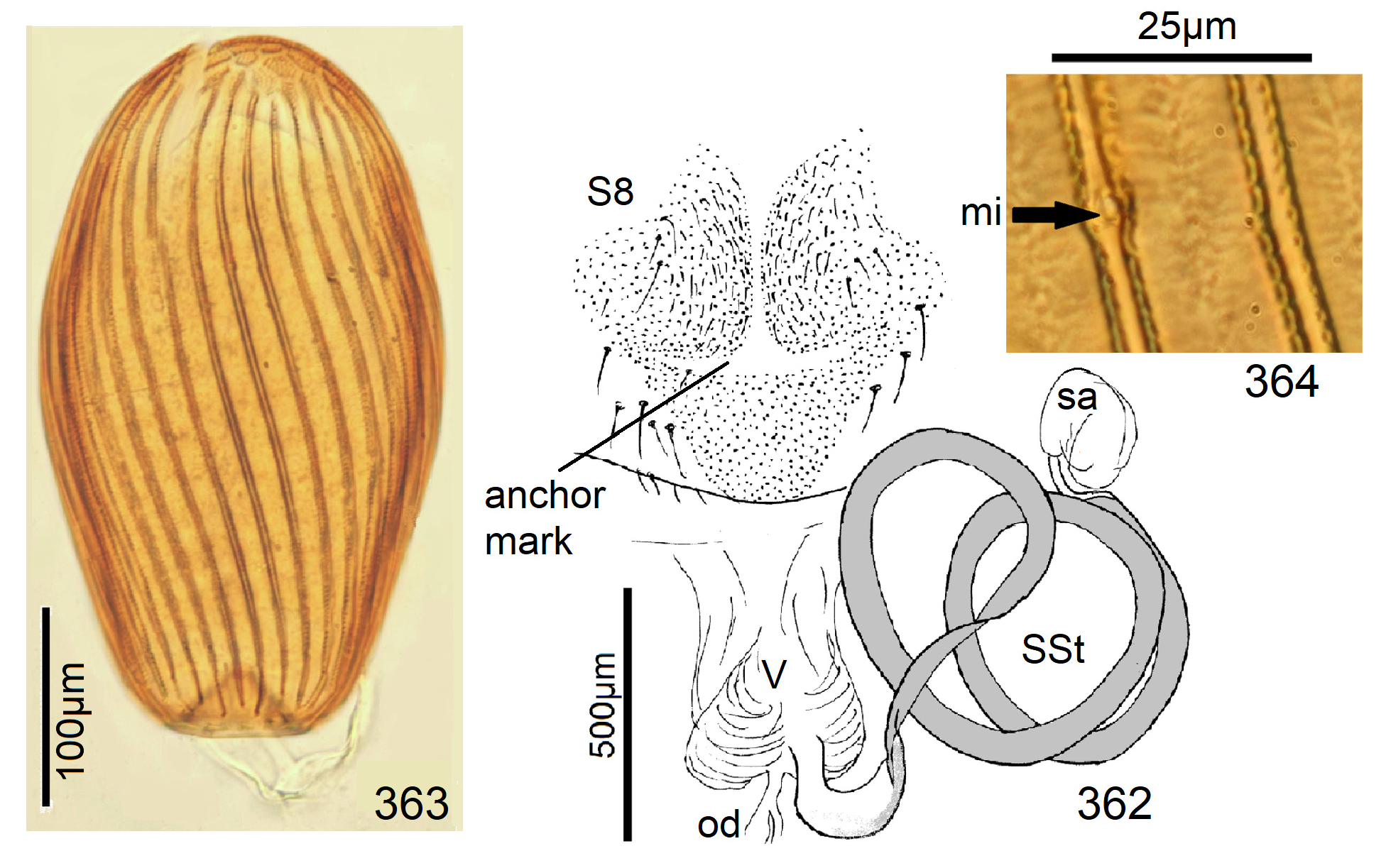
Female Systellognatha share a uniform structure of genitalia. In sternite 7 (S7), the lateral oviducts turn mediad, merge, and form the common oviduct which enters the vagina in S8 on the underside. In preparations, the Y-shaped cuticular oviducal lining is often visible in front of the vagina and the spermatheca (sa) is dorsally attached via a narrow short unmodified spermathecal duct ( SD).
Neoperla View in CoL is the only exception, with several different types of connection between vagina and spermatheca. Three different morphs occurred among the types of Enderlein’s few African Neoperla ( Zwick 1973a) View in CoL , while more were found in Asia ( Zwick 1977). The initial recognition of only two groups ( Zwick 1983) was unfortunate and partly erroneous. However, the distinction of a “ clymene-group ” and a “ montivaga-group ” is still accepted common practice (e.g., DeWalt et al. 2021).
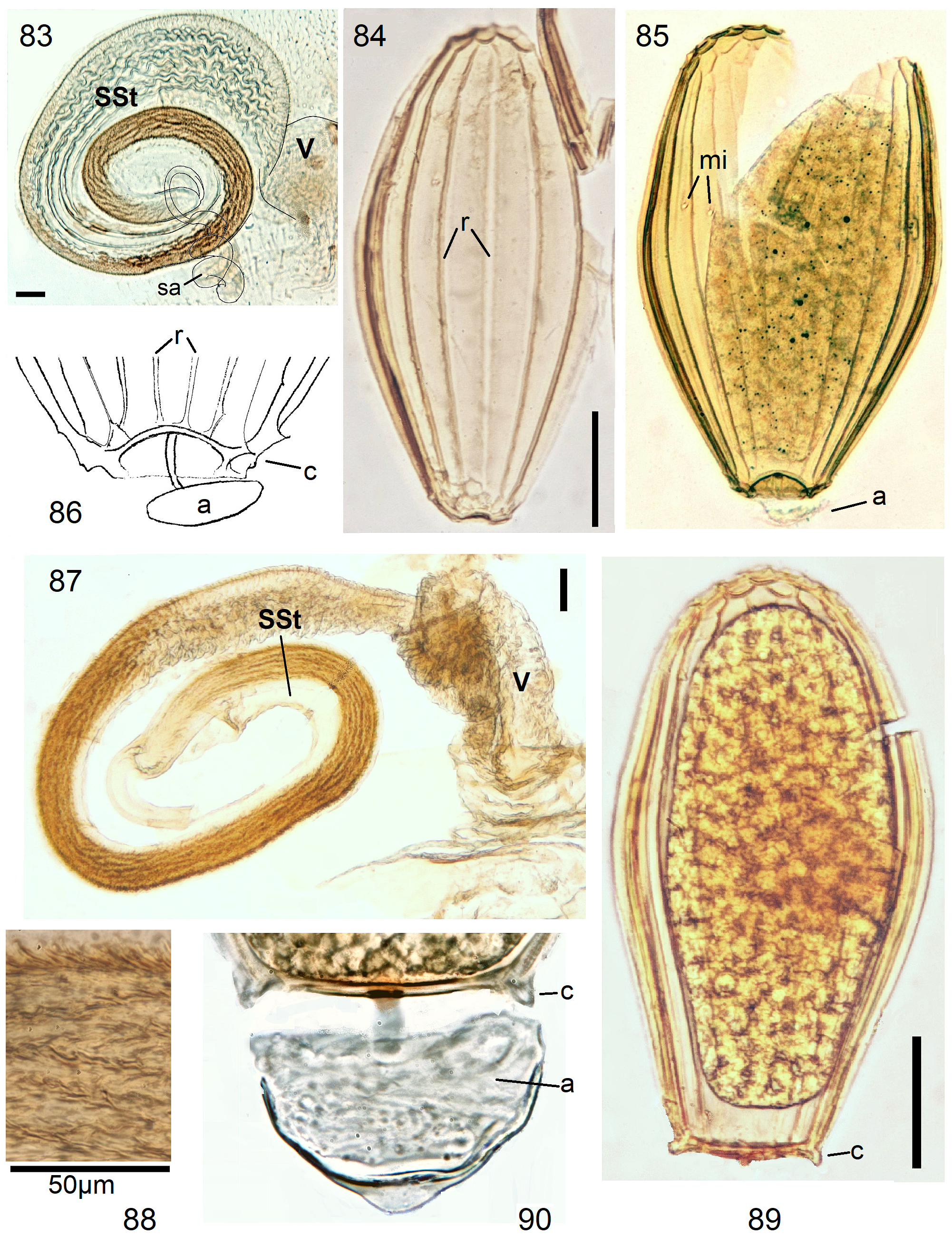
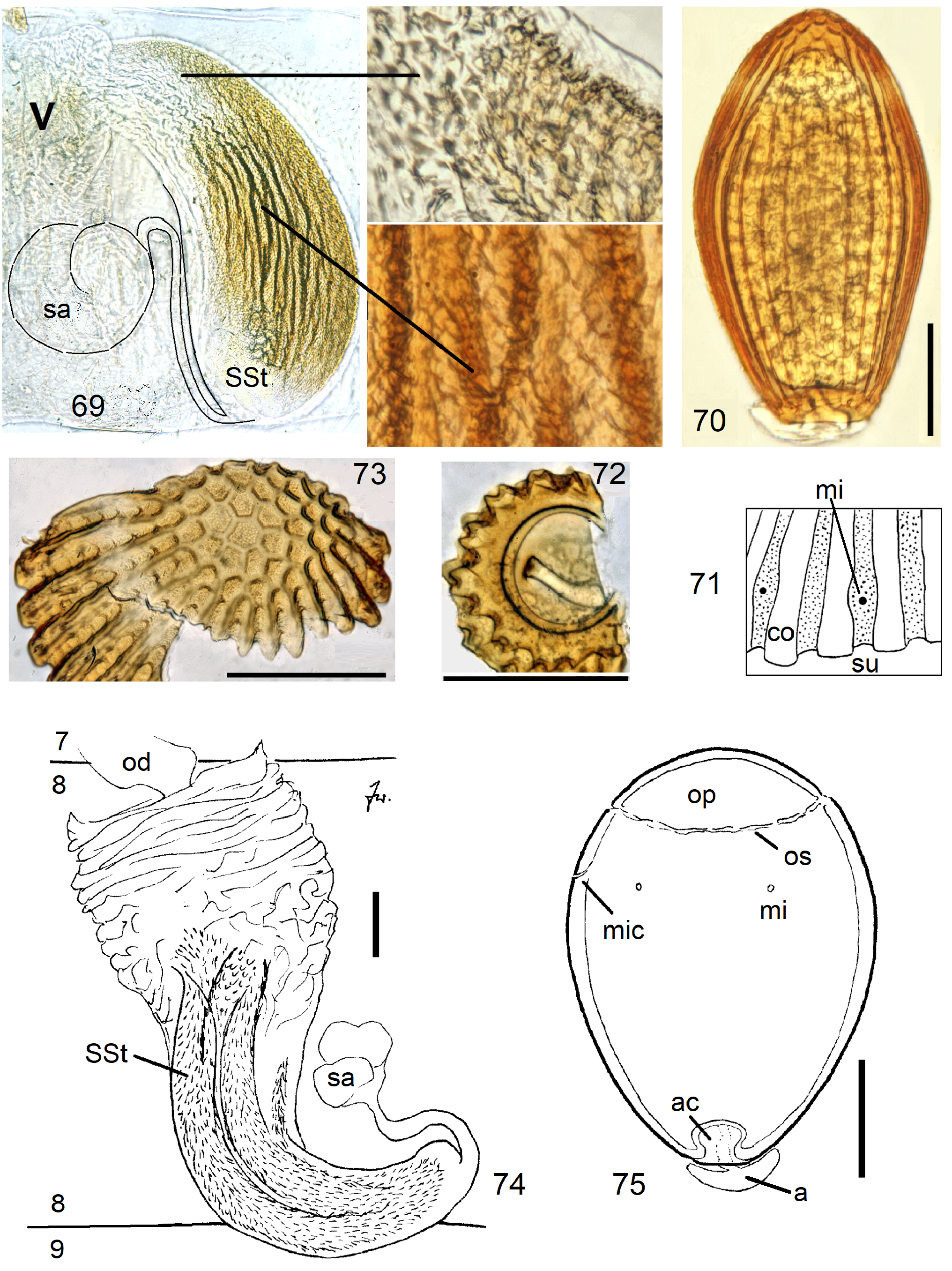
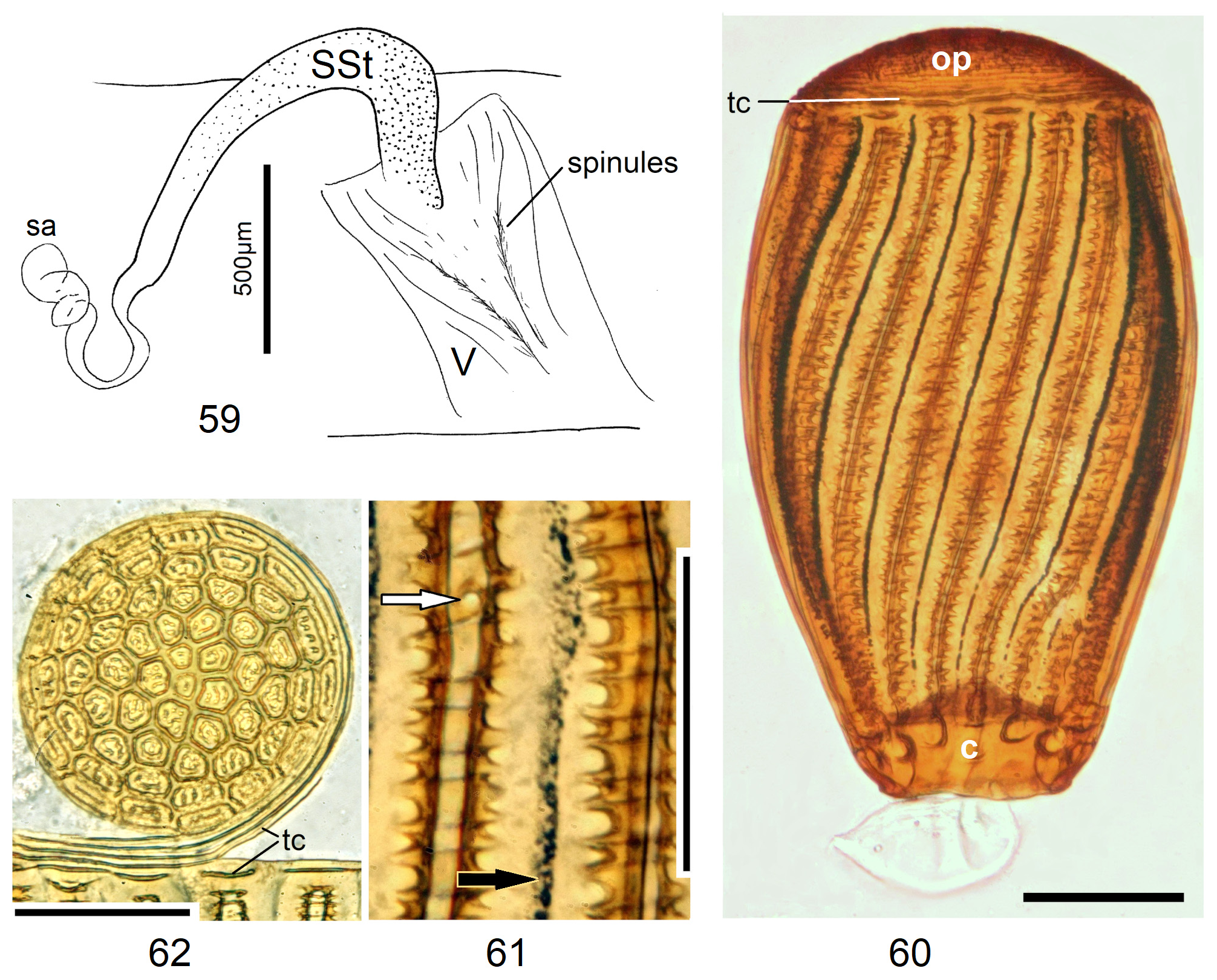
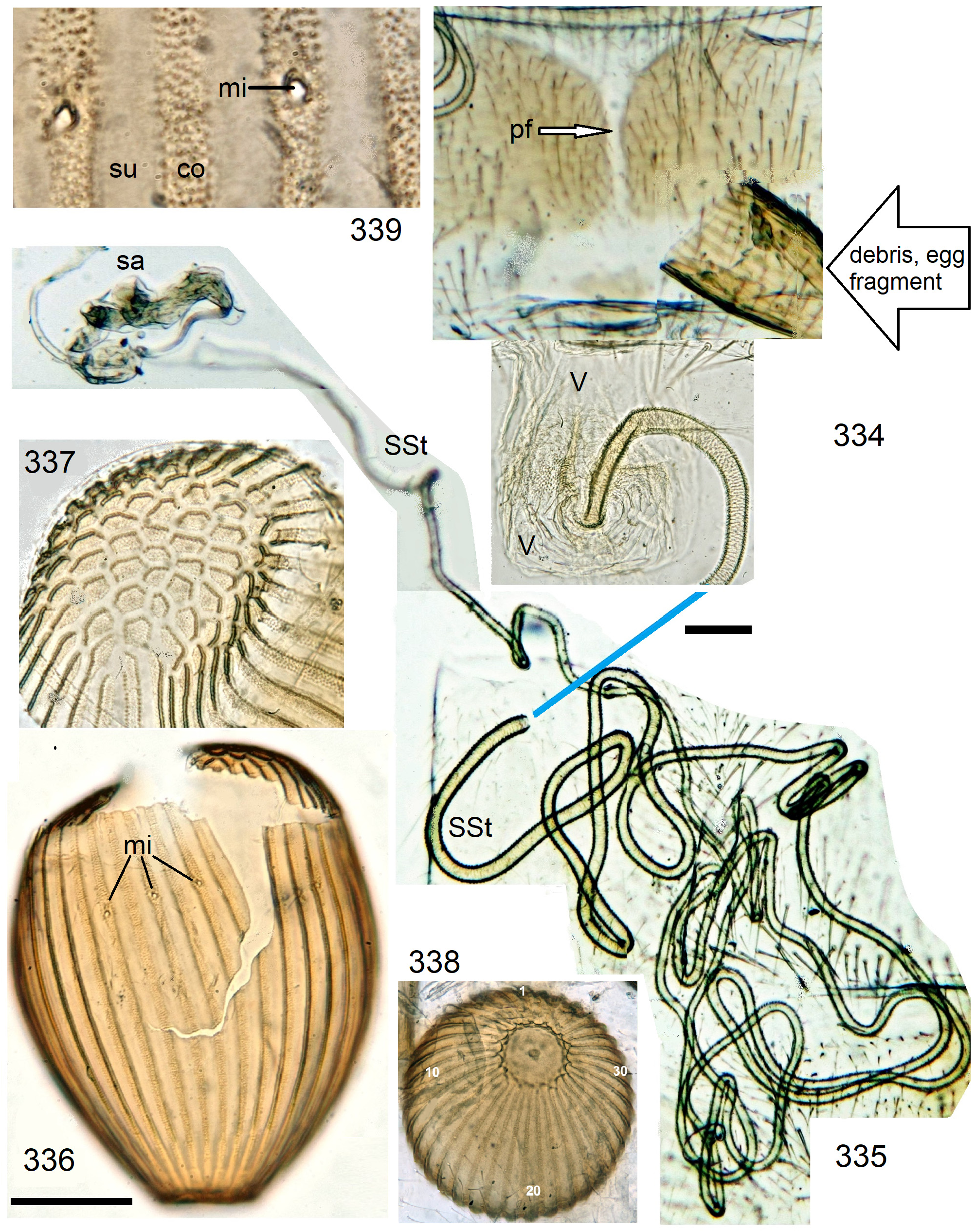
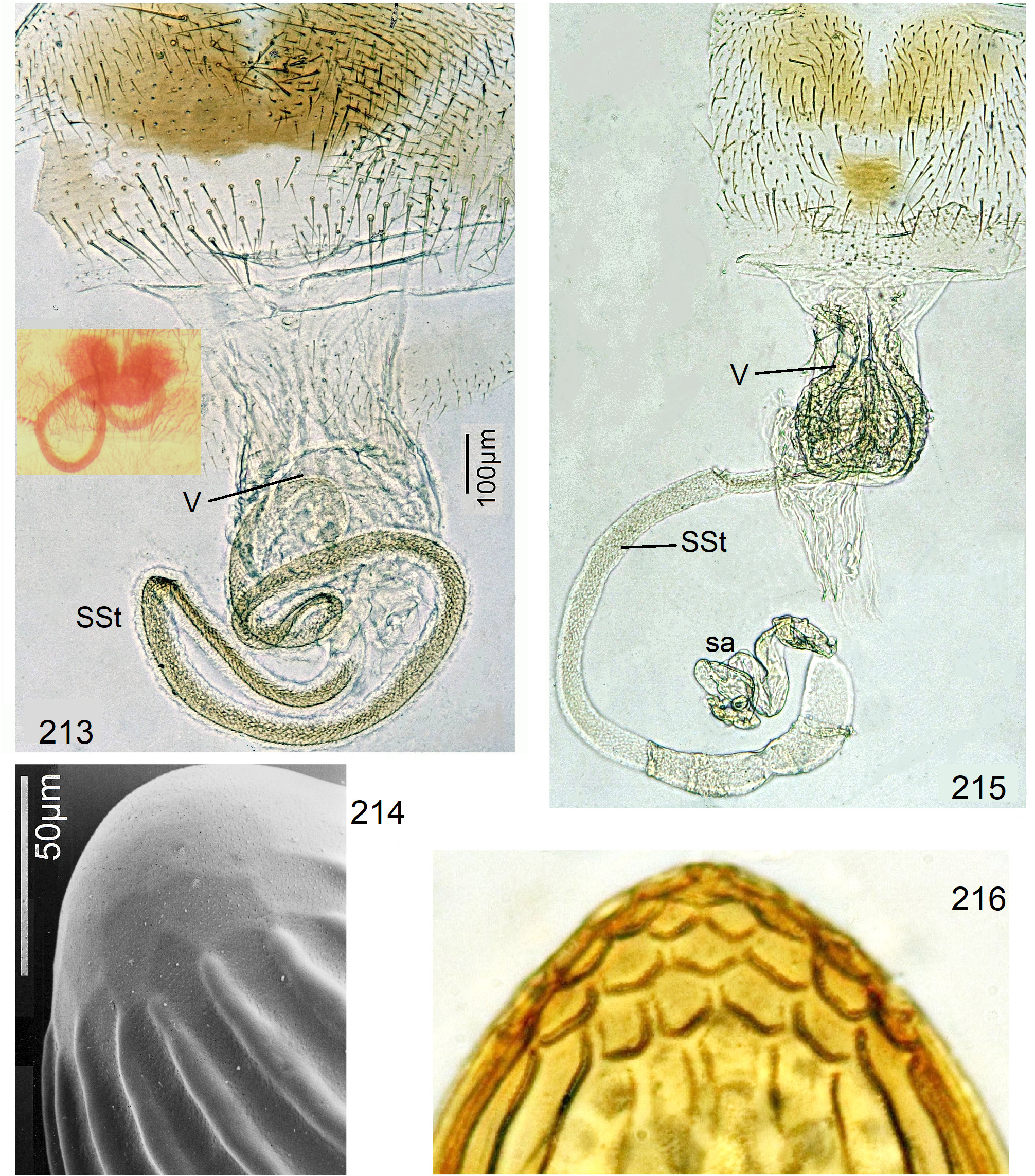
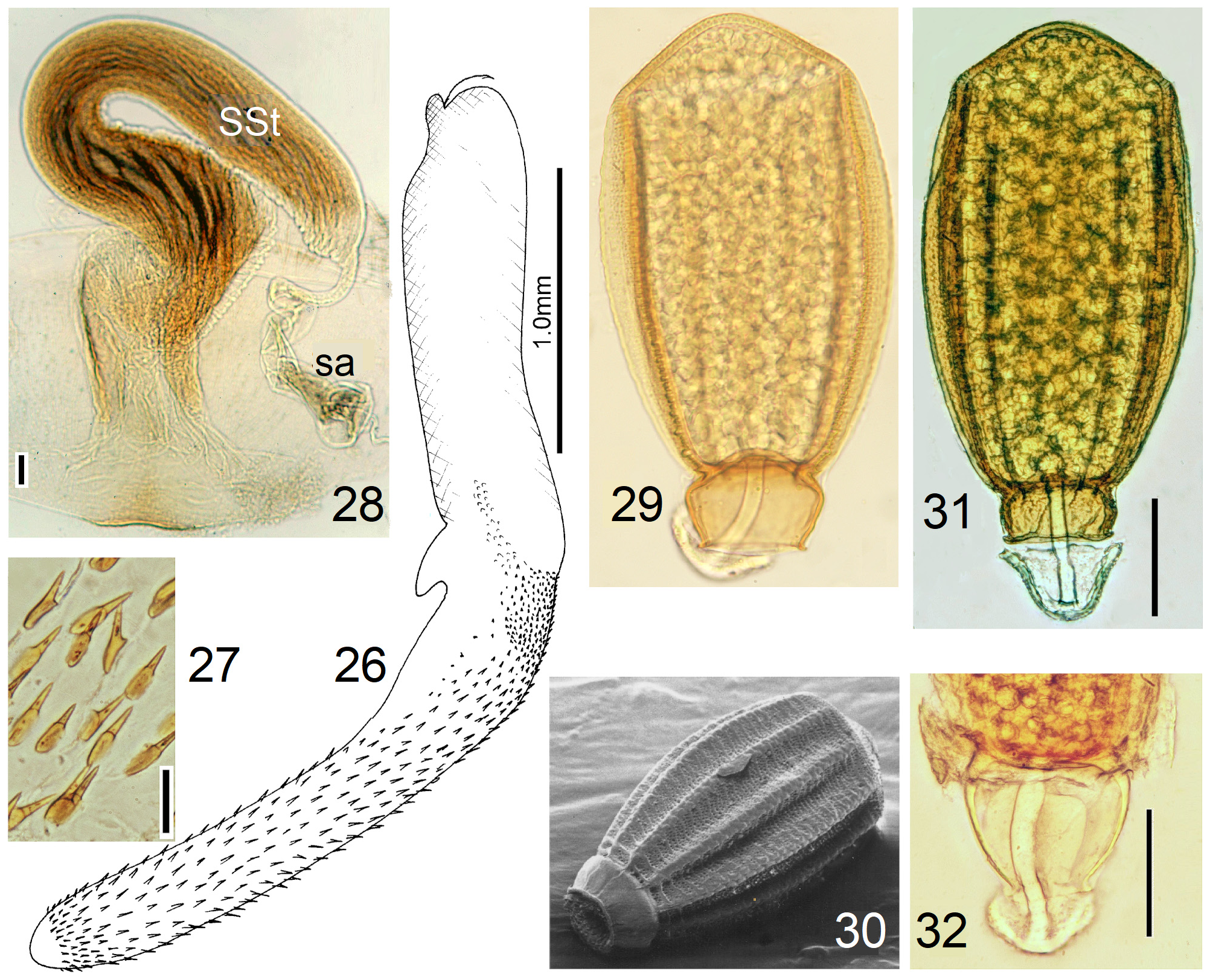
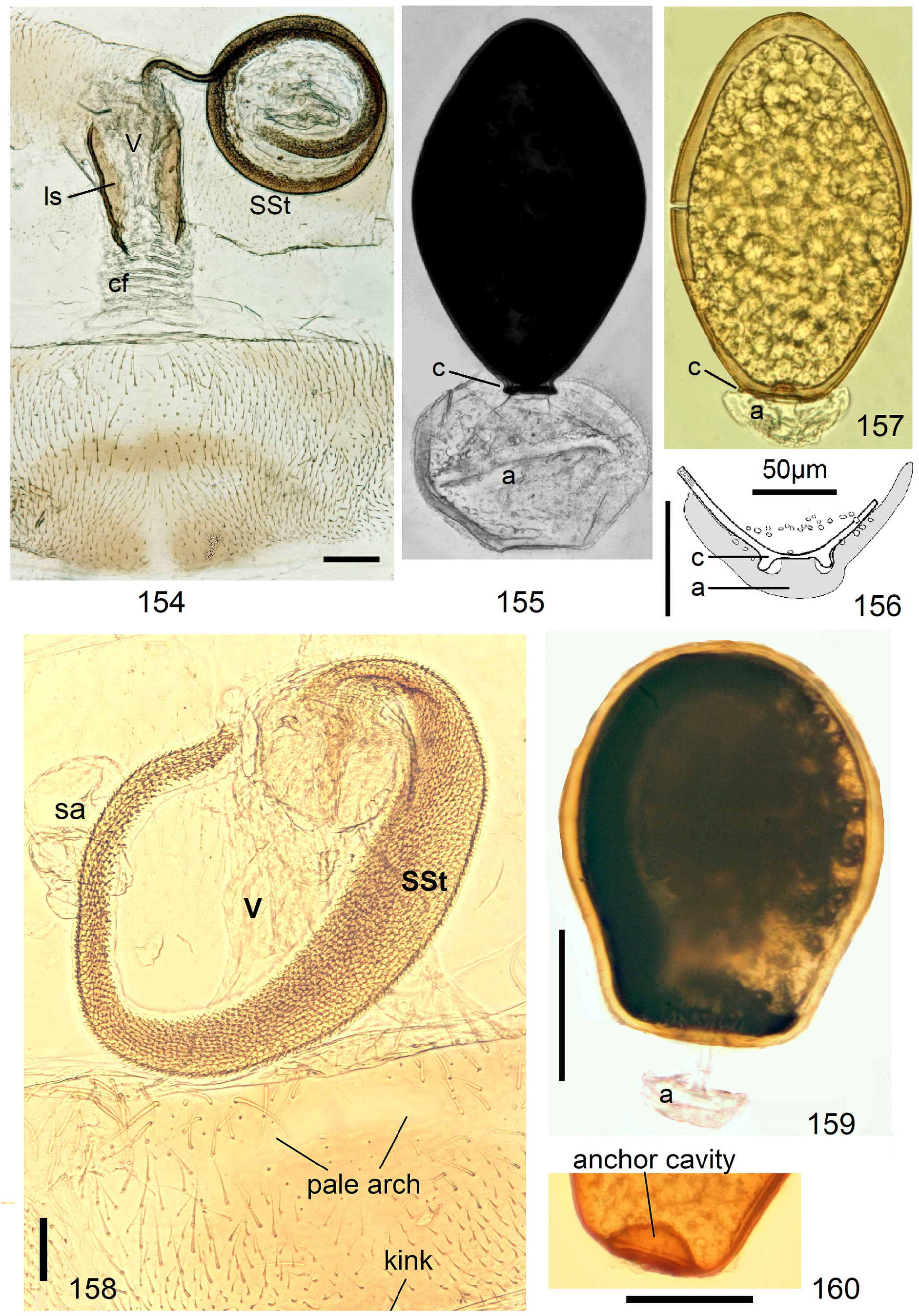
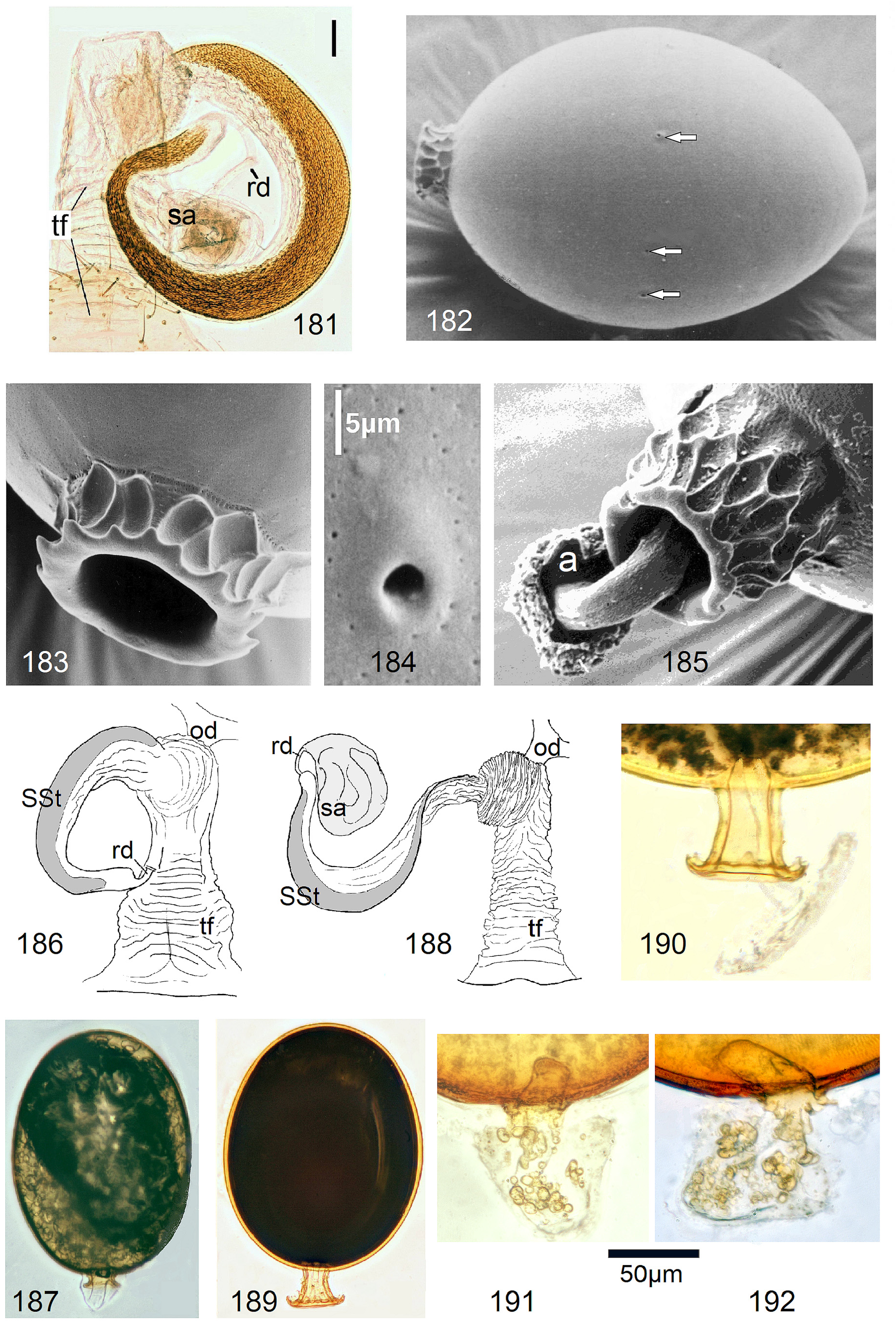
In the forthcoming revised Neoperla View in CoL system (P. Zwick in prep.), all African species are placed in Neoperla (Neoperla) with an armed spermathecal stalk (SSt) connecting the vagina and spermatheca instead of an unmodified cuticular duct. The shape of the stalk and its armature of scales, spinules, and microtrichia differ between groups (e. g., Figs 83, 84 View FIGURES83–90 ). In most African species, the SSt forms a ring or is coiled. The spermatheca itself is a delicate spiral or curled tube (e.g., Fig. 17 View FIGURES 16–17 ) while in most Asian and in the North American species the spermatheca is ball-shaped or resembles a thick sausage with rather firm and often also rough cuticle.
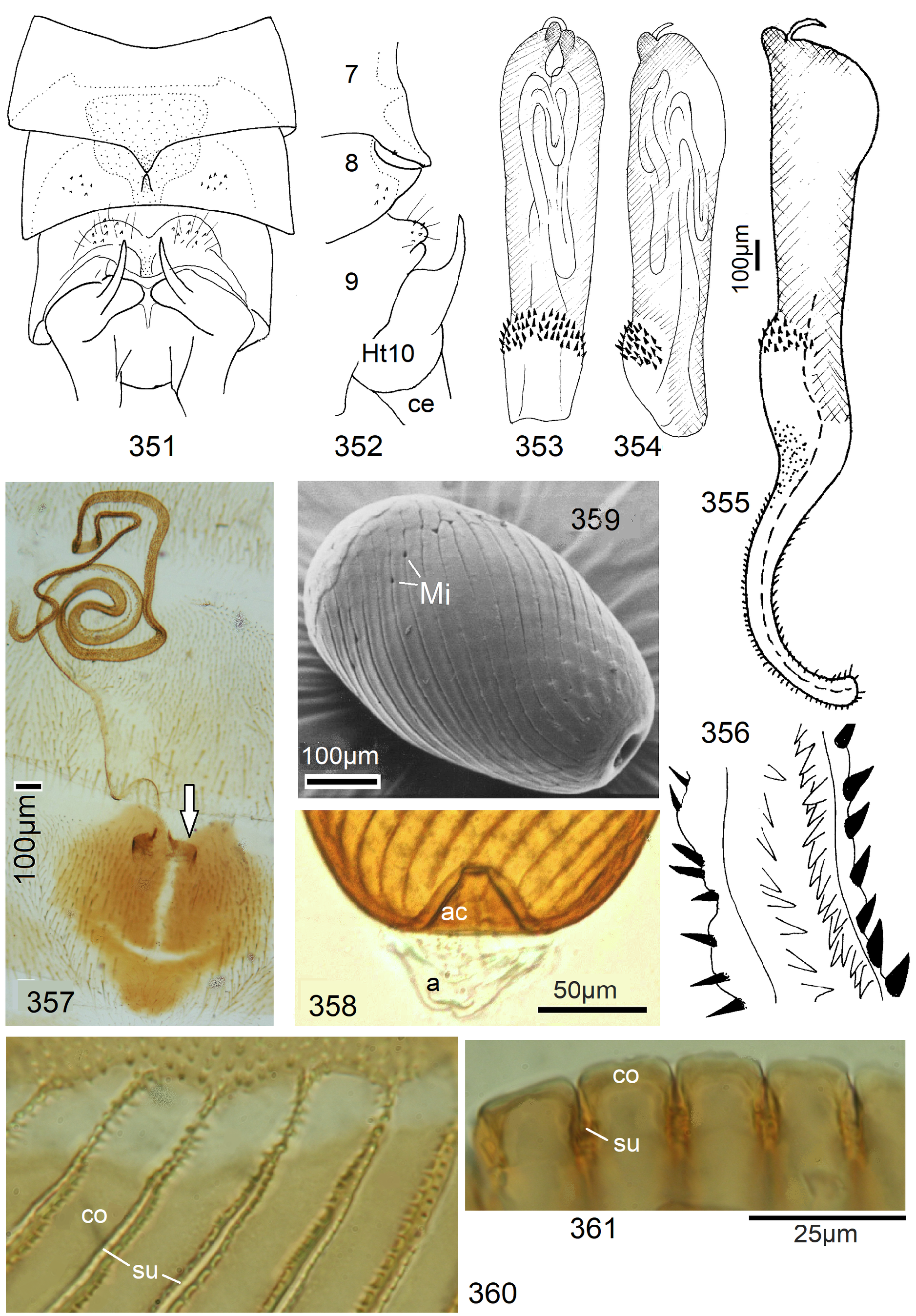
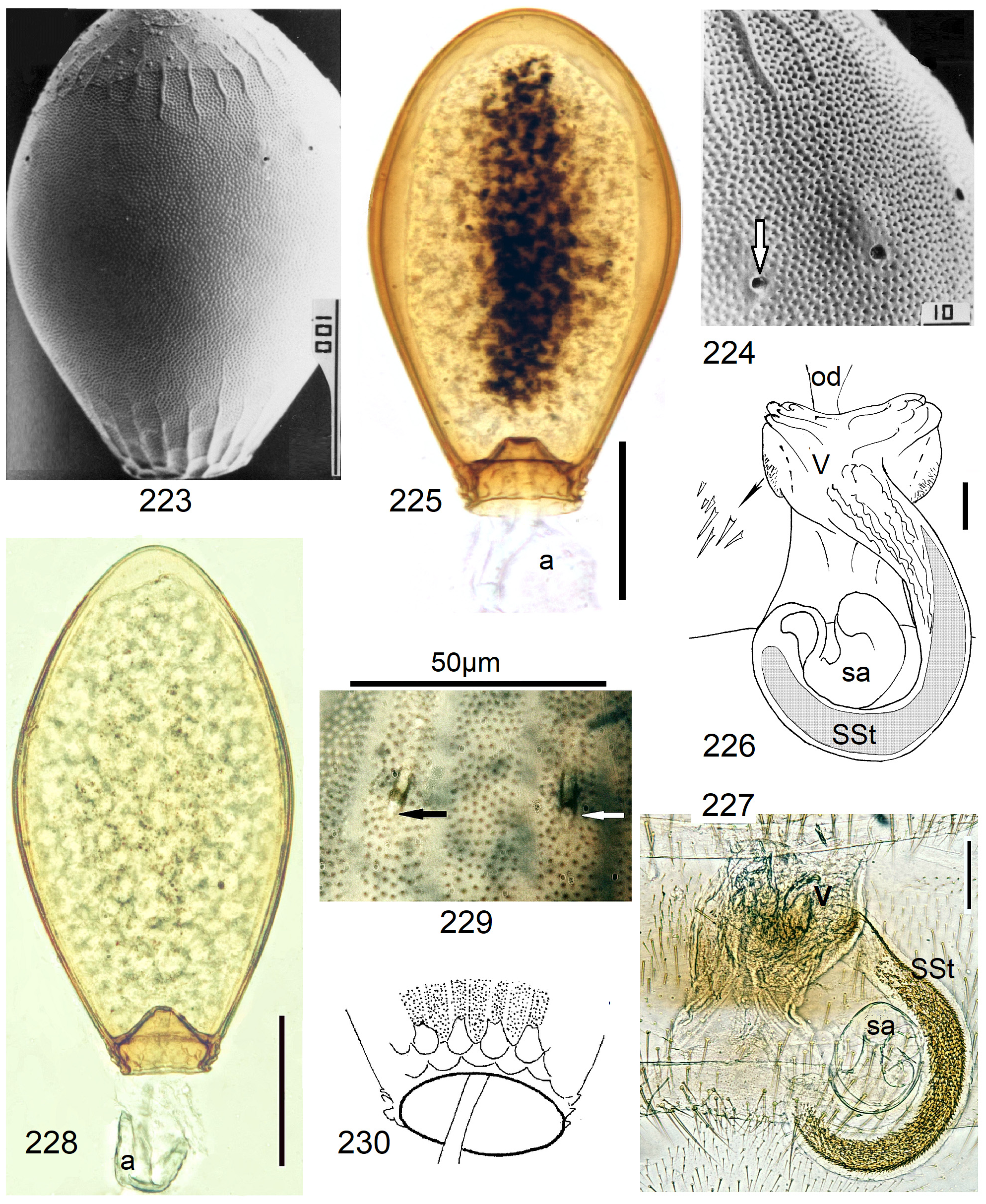
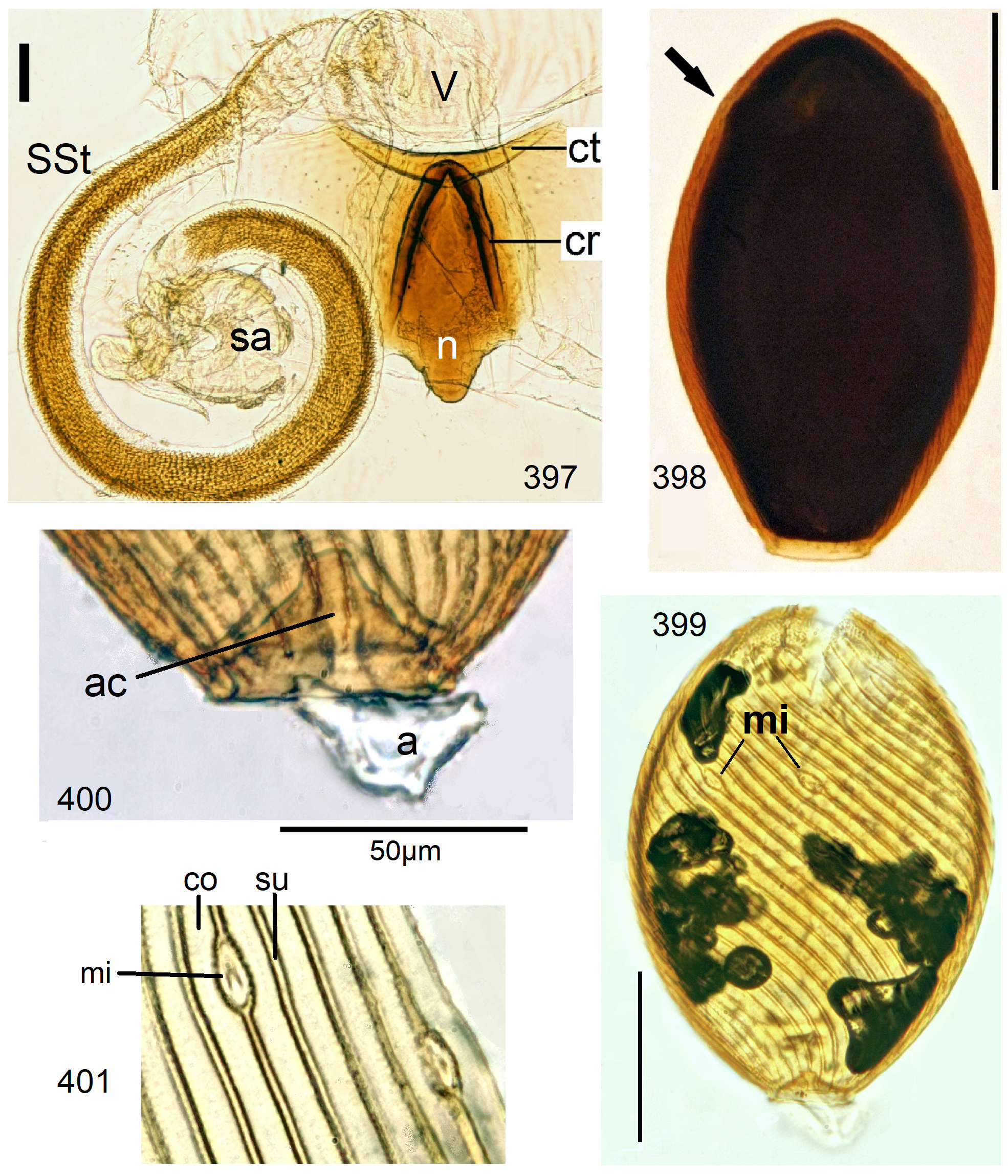
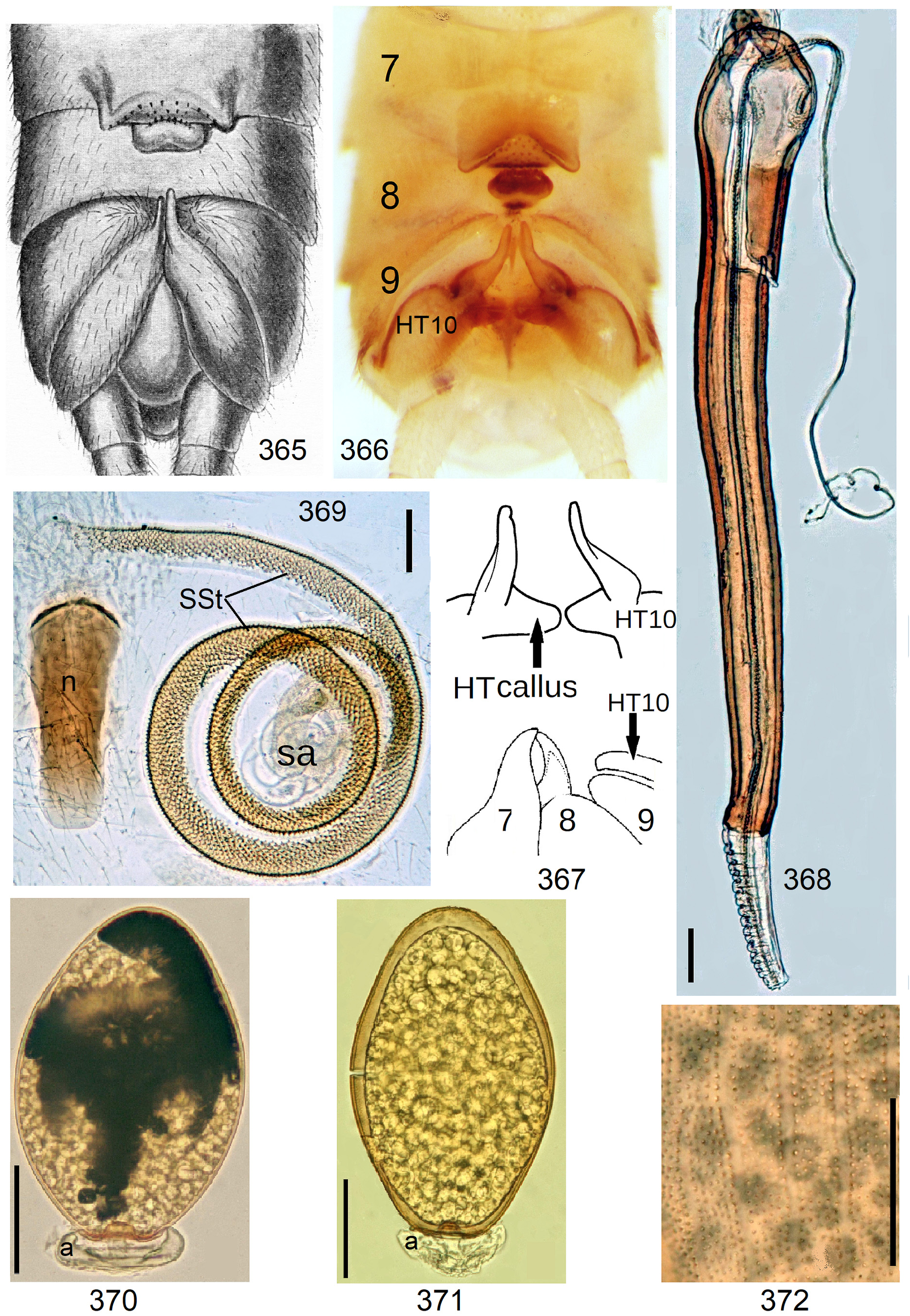
Neoperla View in CoL eggs are ovoid to subspherical; the firm chorion is sculptured by follicle cell impressions ( Rościszewska 1987). Punctures are often called aeropyles (e.g., Picker 1980, Rościszewska 1991, Stark & Szczytko 1988) which seems plausible but in Plecoptera View in CoL there is no evidence of the suggested function. Aeropyles in the sense of Hinton (1981) are different. In the literature, the terms anterior—posterior for the structurally different egg poles vary. We use terms based on function and distinguish the anchor pole from the opposite opercular pole where larvae hatch. The anchor is an elastic attachment structure surrounded by a strongly sclerotised collar; vestigial character expressions and secondary reductions occur. The anchor pole leads the way out of the ovariole into the oviduct when eggs are ready. We use the term operculum for the opposite pole although we do not know if the larva indeed detaches a polar cap or if the larvae exit through some slit in the chorion. However, some African species have a visible opercular suture or a ring of specially shaped follicle cell impressions. Insemination occurs via several micropyles distributed in a ring between widest egg diameter and opercular pole.
Eggs of many Neoperla species are striate, which is unique among Perlidae View in CoL . The plesiomorphic striation is longitudinal ridges on the entirely punctate chorion, with chorion surface differing at best gradually between the ridges and the wide concave interstices between them (e.g., Figs. 85, 87 View FIGURES83–90 ). Many of the species in question have opercula in the shape of a low cap. The apomorphic condition is differentiation of the chorion into alternating raised costae with impunctate usually flat back and punctate concave sulci, with one costa and one sulcus together constituting a stria (e. g., Fig. 86 View FIGURES83–90 ). In most species, striae are straight; spiral striae are apomorphic. The direction of spin is species-specific: striae are called levogyrous if in polar view spirals turn left (anticlockwise), in side view levogyrous striae run from top left to bottom right. Different types of punctation of the sulci are described under the respective species groups.
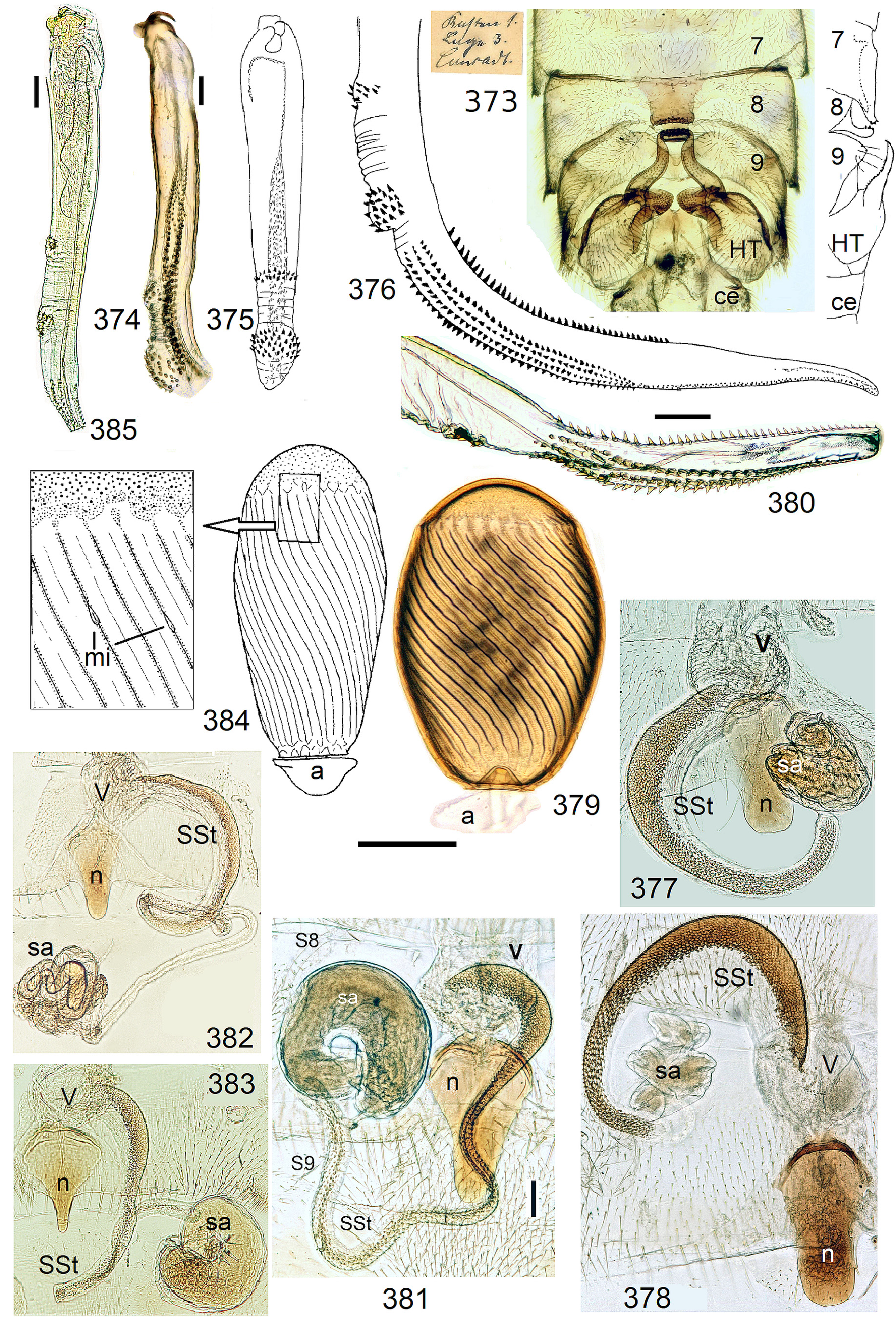
The most derived character expression is a secondary egg surface consisting only of wide bare costae separated by narrow fissures leading to sulci concealed in the depth of the chorion (e.g., Figs. 359 View FIGURES 351–361 , 379 View FIGURES 373–385 ). This is an extreme character expression in a group of species where even freely visible sulci have no punctures and only regular single rows of micropunctures separating sulcus and costa. Micropunctures are structures visible only in mounted eggs at high magnifications, but under transmitted light their structure remained unclear even at magnification 630X.
I. Keys to species groups and species of African Neoperla (Neoperla)
Dichotomous keys to taxa at various levels (e.g., species co-complexes) and to all species are provided. However, only one gender is known for several species. To be user-friendly, some species appear at several places in a given key or in several keys.
All characters used in a key couplet must be considered and identifications should be checked against the individual descriptions and illustrations. Specific identification of females without eggs may be impossible.
Males
1 T7 with an unpaired caudo-medial process; T8 variable....................................................... 2
1’ T7 without unpaired caudal process; T8 with a forward-curved process or hook........................................................................................... N. excisa View in CoL and N. sjostedti View in CoL -operational complex, p. 130
2 Caudal process of T7 flat (e.g., Figs. 1, 4 View FIGURES 1–5 , 11 View FIGURES 11–15 ); T8 not or only weakly sclerotised along midline, no humps or processes; penis soft, only basally sclerotised and, when contracted, short and flat; the endophallic armature is dispersed, dorsal and ventral sides are not much different (e.g., Figs. 2–3 View FIGURES 1–5 , 12 View FIGURES 11–15 )......................................... N. transvaalensis View in CoL -group, p. 19
2' Caudal process of T7 variable but always three-dimensional, not flat; T8 distinctly sclerotised in midline (e.g., Figs. 76–77 View FIGURES 76–82 ); penis tubular, sclerotised, endophallus variable.............................................................. 3
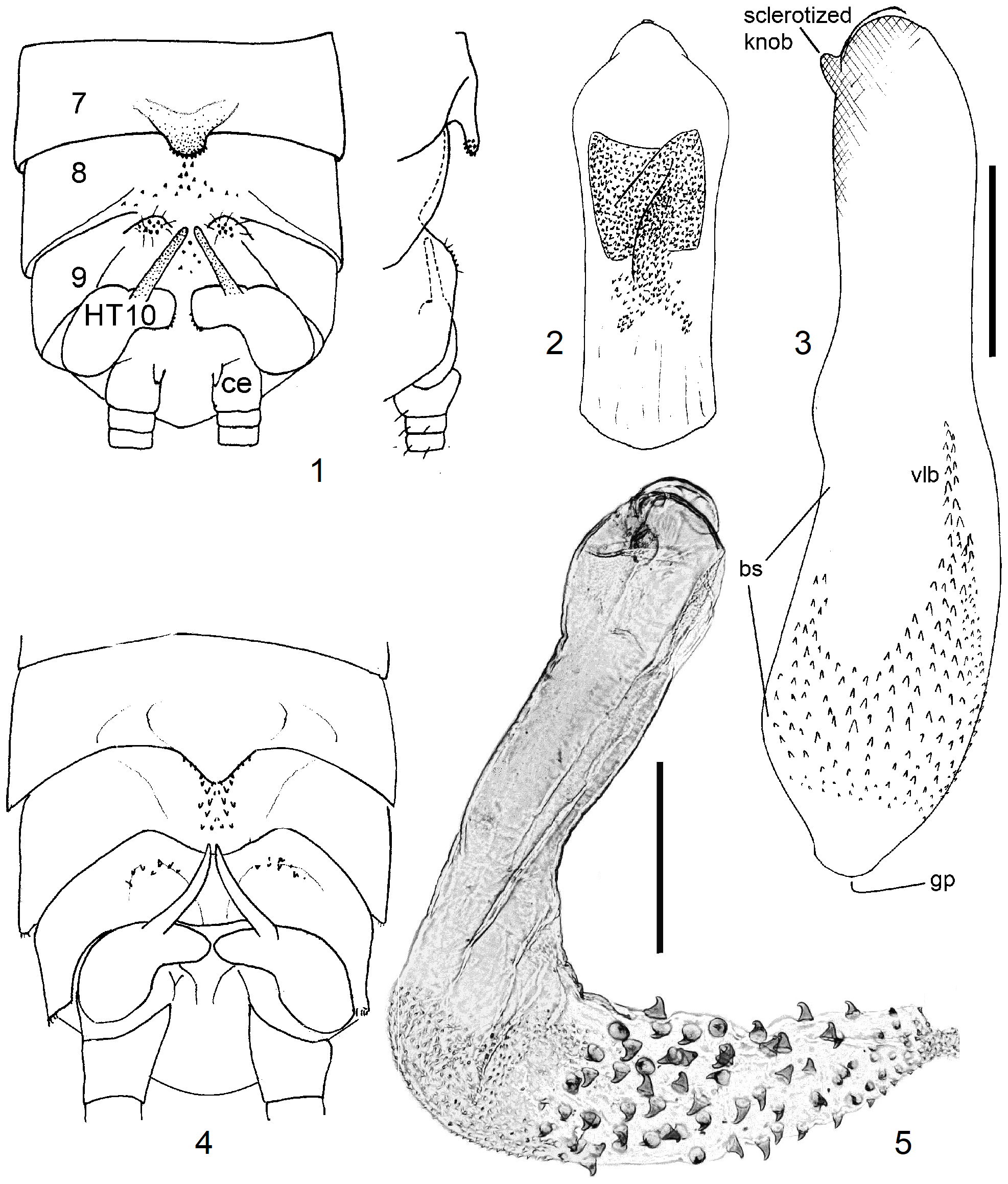
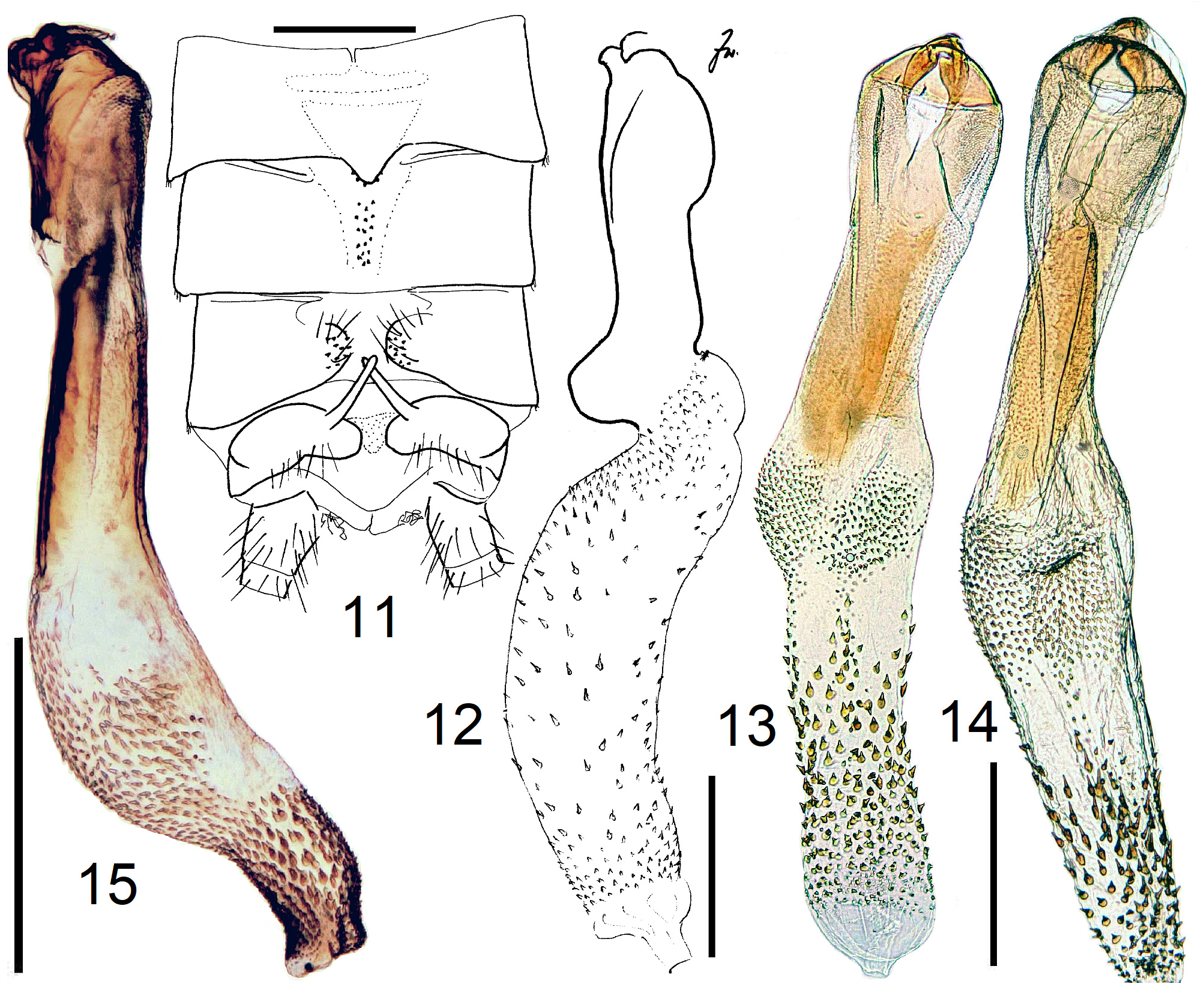
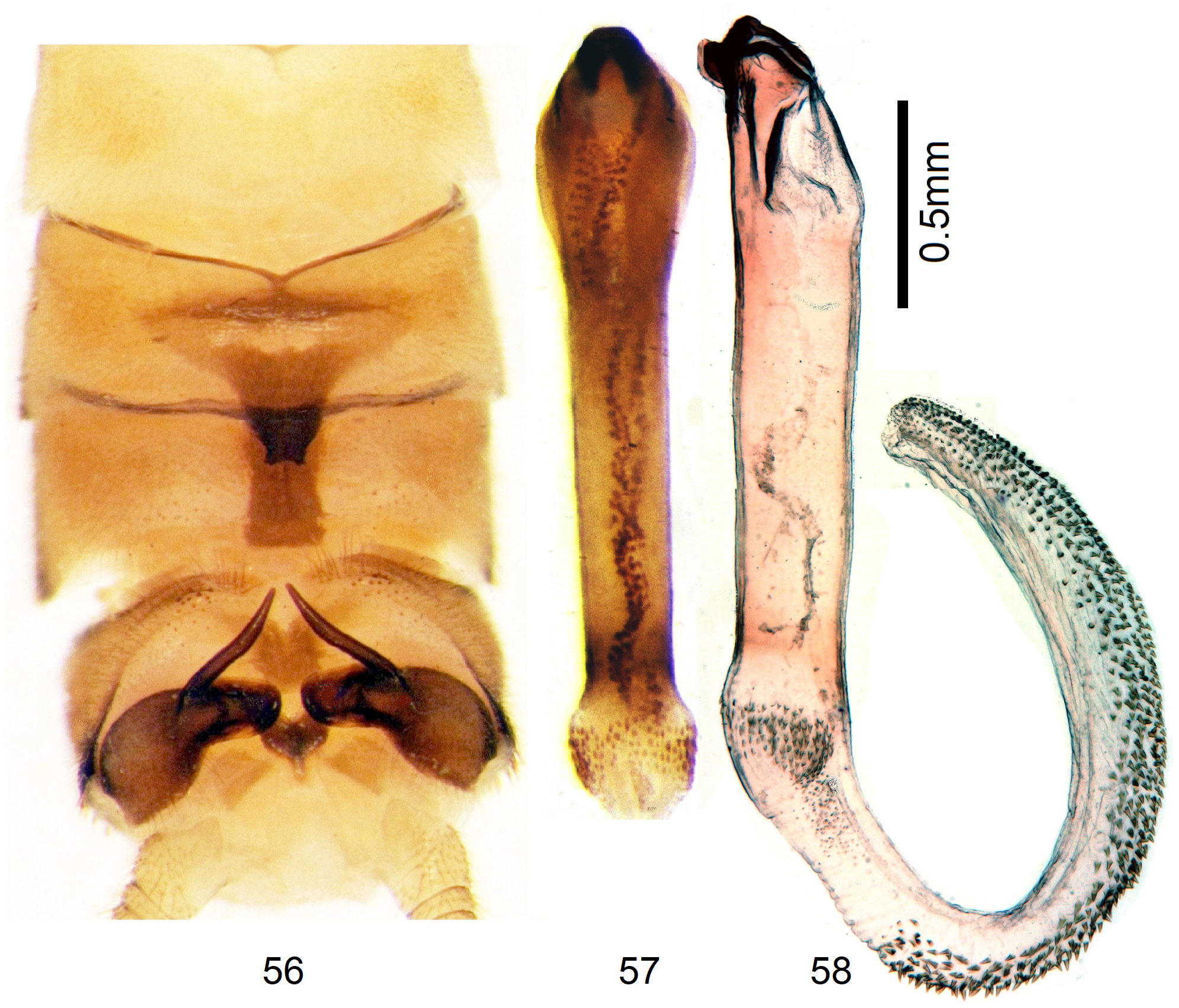
3 Process of T7 distally wide and with a pair of short, small tips; sclerite on T8 band-shaped. Hemitergal process long and sharply pointed. Endophallus curved, concave side bare, convex side with dense cover of small spines ( Figs. 56–58 View FIGURES 56–58 )................................................................................................ 11 N. ivanloebli n. sp.
3' Process of T7 with single tip, remainder variable............................................................ 4
4 Process of T7 slightly downcuved, parallel to the caudally rising surface of T8; HT10 with long sinuous tip, mediobasal callus large, square, the wide endophallus as long as the penis, straight or curved ( Figs. 91–93 View FIGURES 91–95 ).......... 19 N. planidorsum n. sp.
4' Process of T7 not parallel to the surface of T8, more or less raised............................................... 5
5 Process of T7 narrow, sharply pointed and very long, extending over most of T8................................... 6
5' Process of T7 not unusually long, extending only a short distance over T8, variable................................. 7
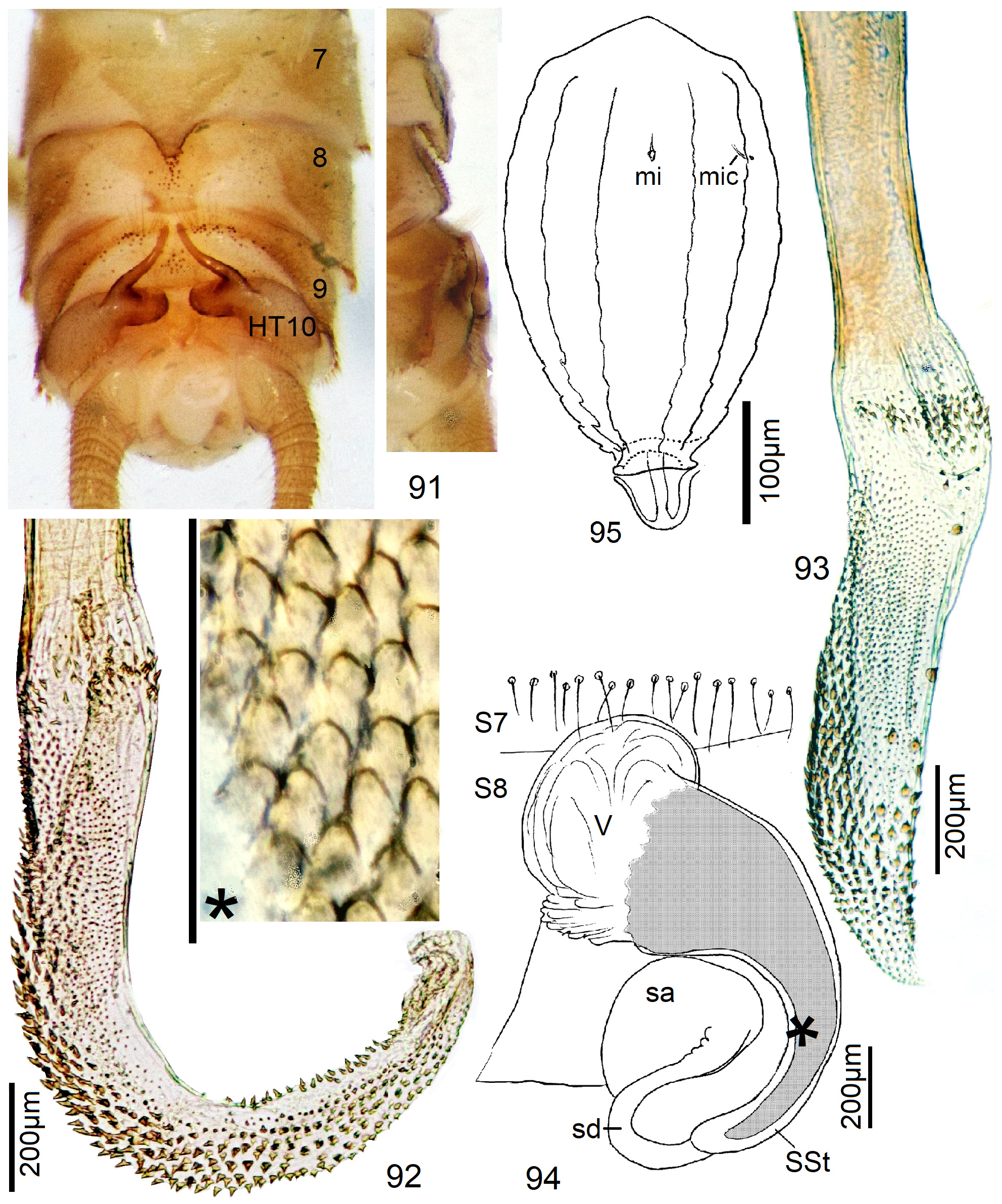
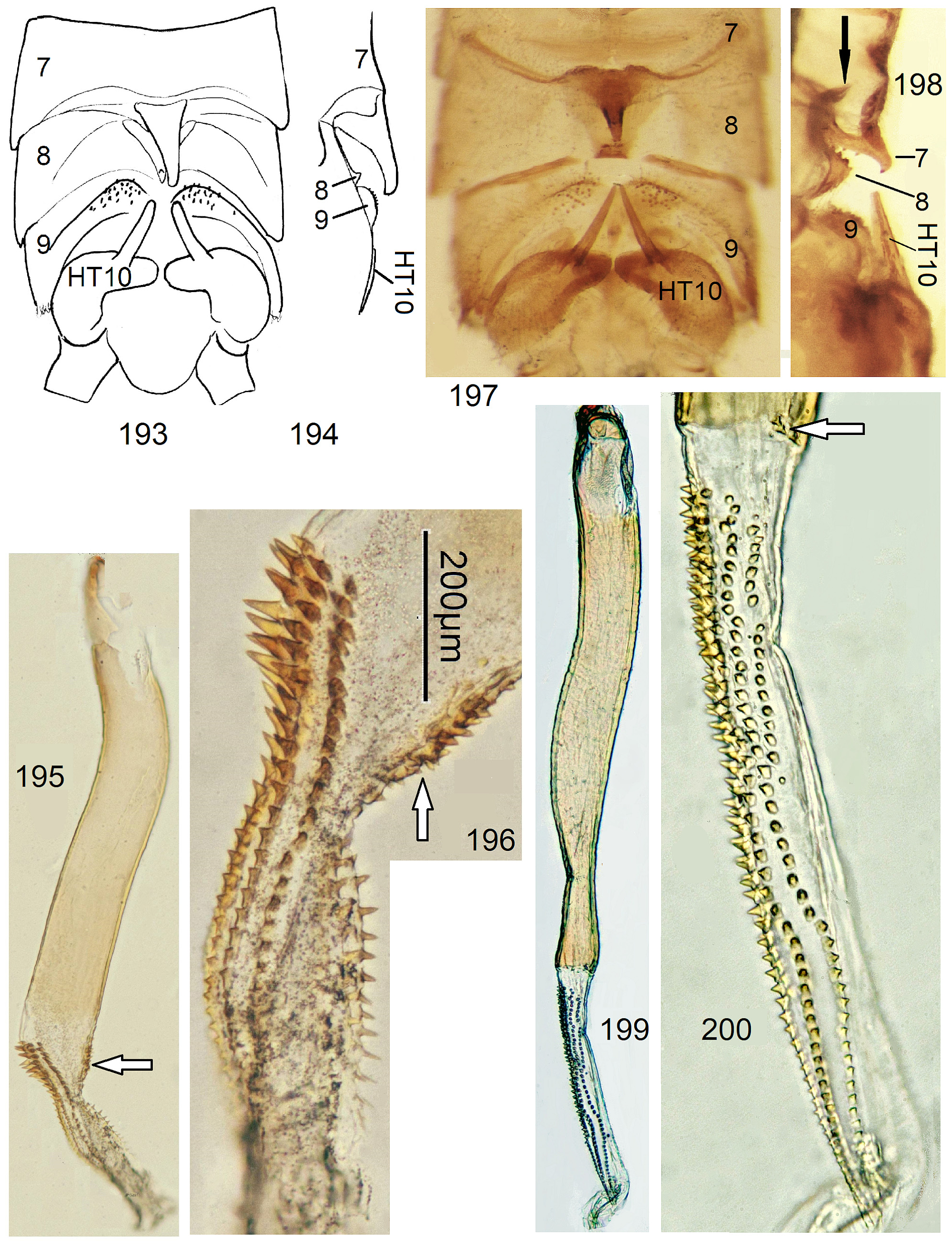
6 Process on T8 minute, penis angled near middle, ventrodistally with external spine patch, the short endophallus dorsally with approximately 7 rows of large spines, ventrally a single row of slender spines ( Figs. 193–196 View FIGURES 193–200 ).......... 35 N. pusilla n. sp.
6' A strong hump with forward skew on T8, HT10 with straight sharply pointed tip, endophallus nearly half as long as penis, dorsally with long regular spine rows, ventrally bare ( Figs. 197–200 View FIGURES 193–200 )......................... 36 N. multiserrata n. sp.
7 Process of T7 a short erect knob, T8 bears a pointed erect process. Hemitergal process slender and long, with modified tip. Endophallus shorter than penis tube, wide, curved ( Figs. 313–322 View FIGURES 313–322 ).......................... N. dubia -complex, p. 122
7' Process on T7 longer, directed upward or backward, process on T8 variable....................................... 8
8 T8 with small longitudinal caudal sclerite which can be uplifted to project as a kidney-shaped structure from a slit in the tergite ( Figs. 206–207 View FIGURES 205–208 ).......................................................... 38 N. View in CoL burgeoni and 39 N. beta n. sp.
8' Sclerite on T8 not moveable, shape varies, relief varies from flat to high humps.................................... 9
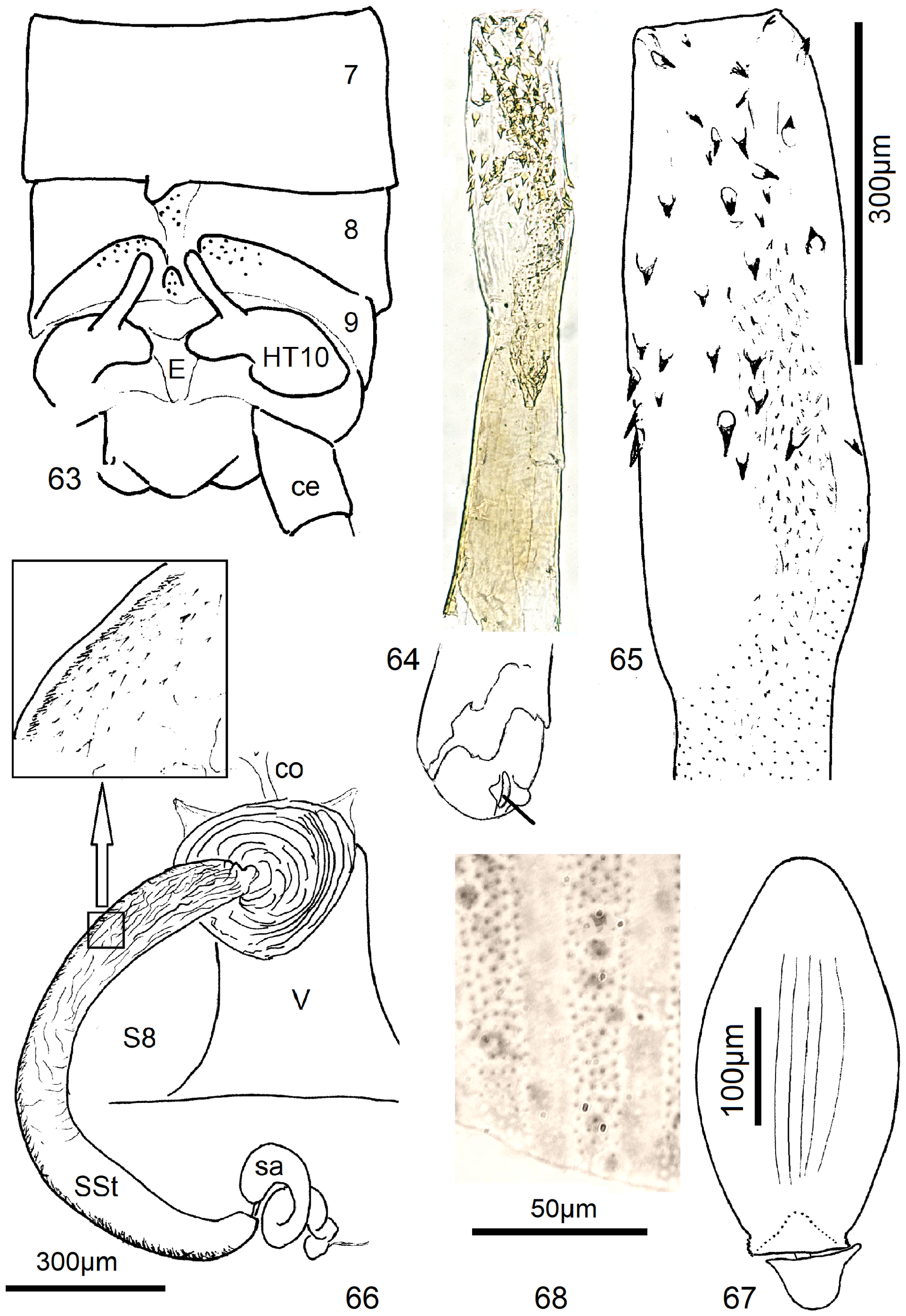
9 T8 with a flat, weak sclerite. A small erect scale seen caudally is present in the median furrow of T9. HT10-process is a short straight finger; armature of endophallus consisting of small spines ( Figs. 63–65 View FIGURES 63–68 ).................. 13 N. angolana n. sp.
9' Sclerite on T8 not flat, with a variable hump, no erect scale on T9, remainder variable.............................. 10
10 Process of T7 relatively long and blunt-tipped, in side view raised obliquely over a strong hump on T8, HT10-process angularly bent. Penis tube and endophallus of similar length, endophallus almost straight, armature not densely packed, ventral face largely bare, dorsodistally a few short spine rows are present ( Figs. 76–80 View FIGURES 76–82 )..................... 16 N. duodeviginti n. sp.
10' Process of T7 is a pyramidal cone, sclerite on T8 variably elevated, HT10 curved................................. 11
11 Penis tube and endophallus of similar length, endophallus almost straight, with spines spreading in all directions, some are large and hook-shaped ( Figs. 102–107 View FIGURES 102–107 )....................................................... 22 N. erinaceus n. sp.
11' Length relation between penis tube and endophallus variable but endophallus curved backward to ventral side, armature on dorsal and ventral sides separate, different................................................................. 12
12 Penis tube and endophallus of similar width, endophallus distally not very narrow and pipe-like...................... 13
12' Endophallus much narrower than penis tube, distal part pipe-like and with very long rows of minute spines, often only 2–3 rows which are visible also in the non-everted condition (e.g., Fig. 240 View FIGURES 238–243 )............................................. 14
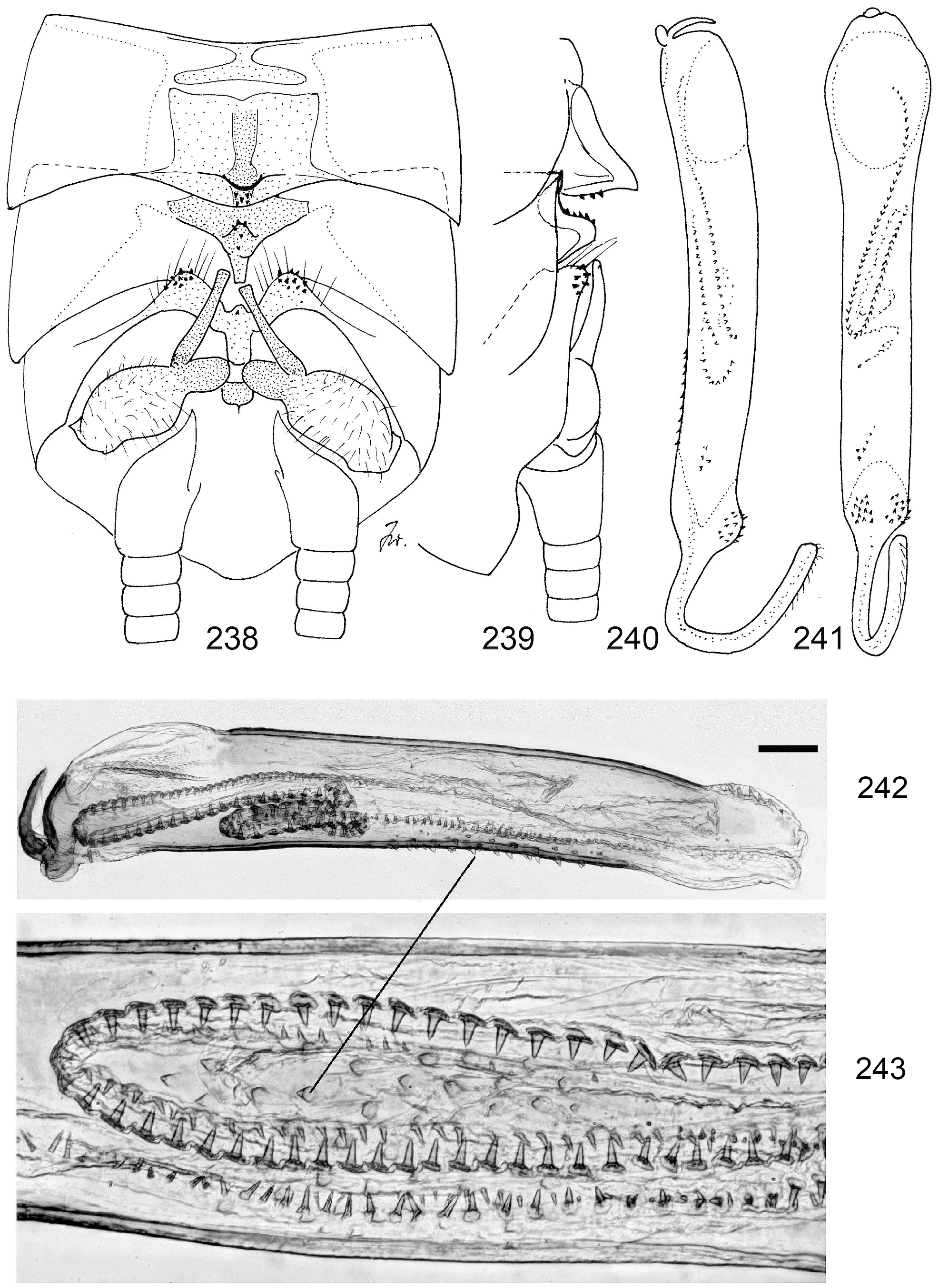
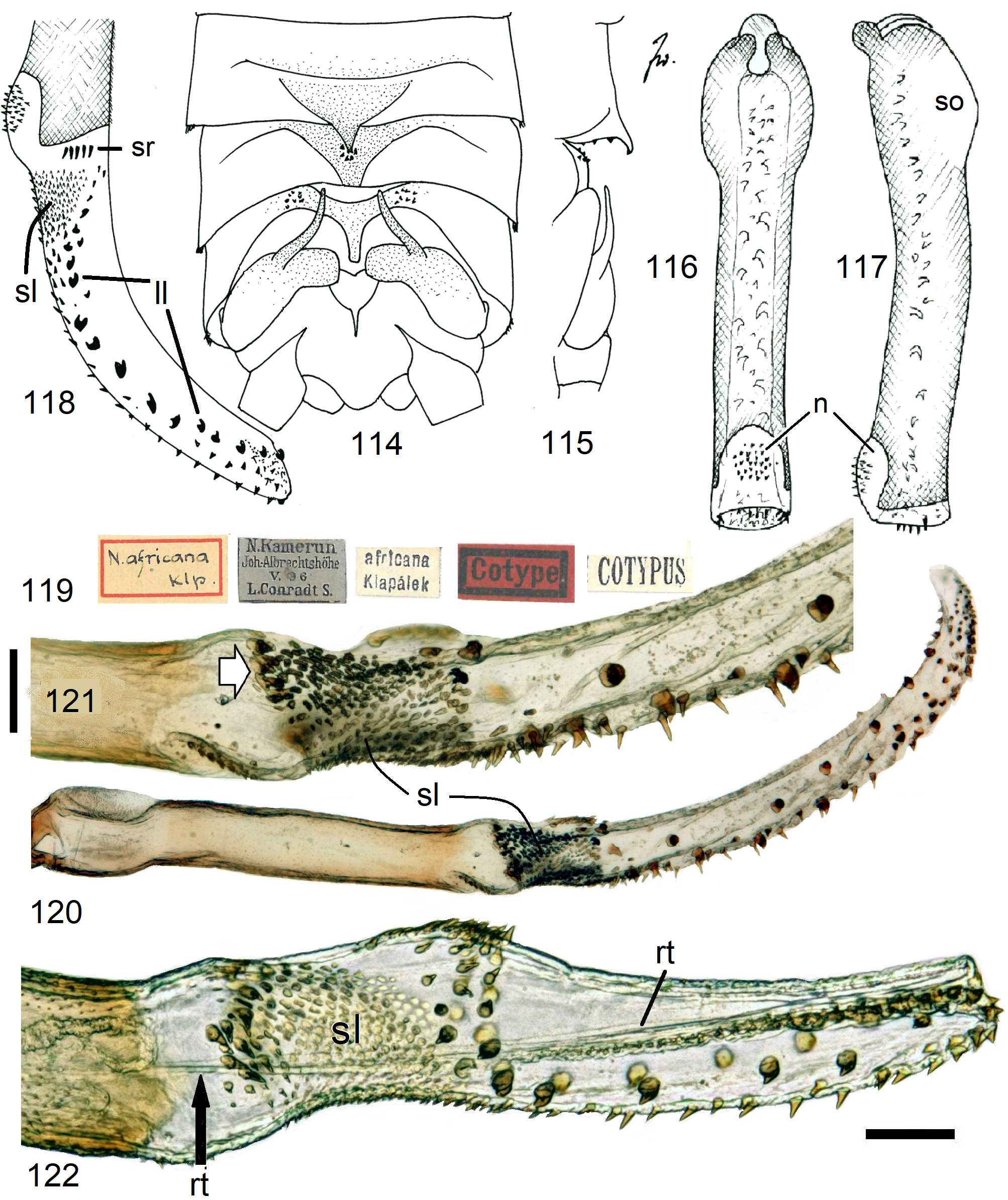
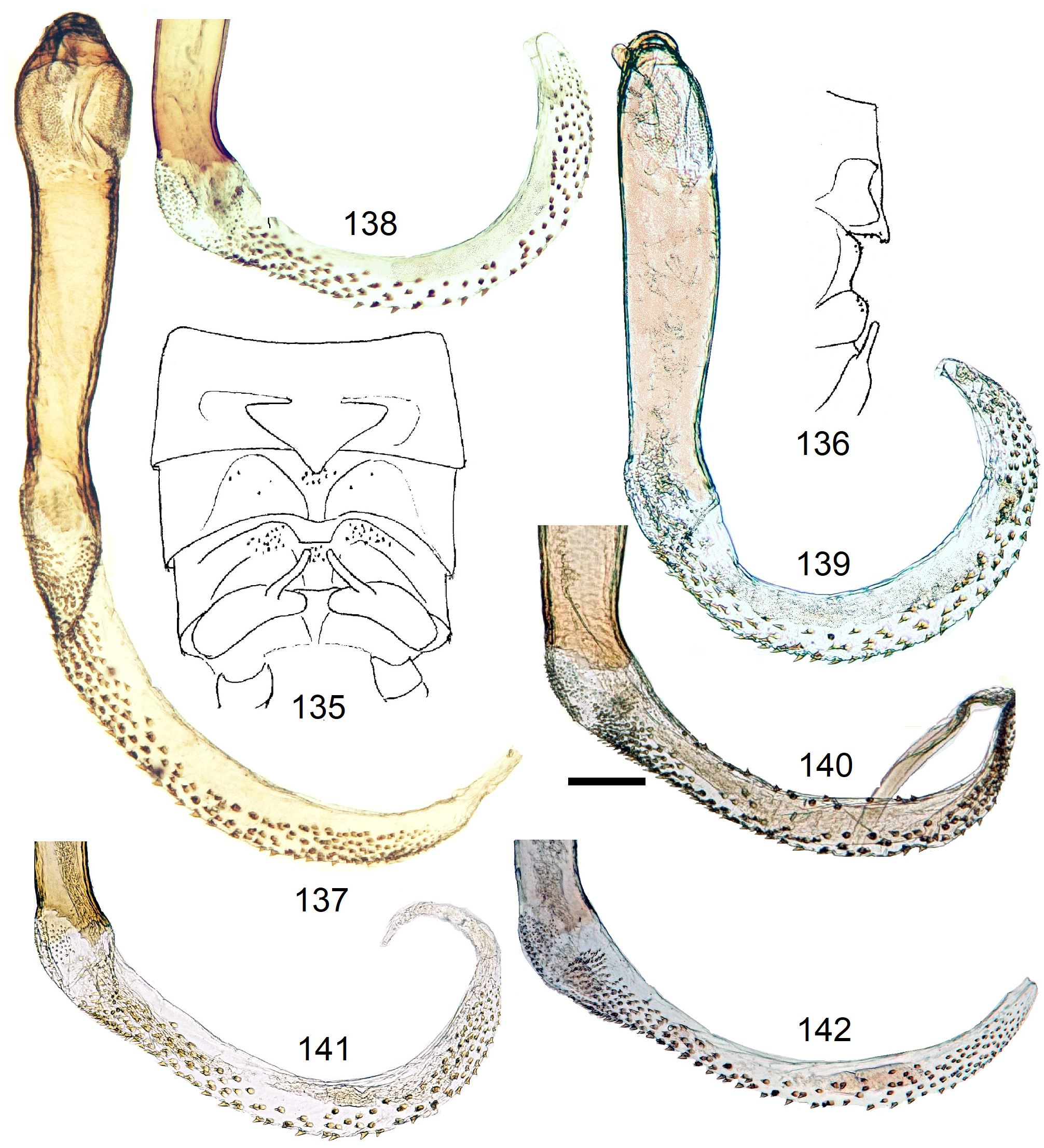
13 Armature begins at base of endophallus, right beyond penis tip (e.g., Figs. 118 View FIGURES 114–122 , 137 View FIGURES 135–142 )............. N. africana View in CoL -group, p. 50
13' A bare section between penis tip and armature on endophallus (e.g., Fig. 164 View FIGURES 161–168 )............... N. pilulifera -complex, p. 70
14 Penis tube with dorsolateral, dorsodistal, and ventral patches of coarse external spines ( Figs. 296–300 View FIGURES 294–304 )................................................................................................... 52 N. orthonema n. sp.
14' Penis tube without patches of external spines, rarely a few single external spines................. N. spio View in CoL -complex, p. 87
Females
1 S8 with a large caudal notch, a projecting nail, or a median sclerite resembling a fingernail. SSt coiled, inside with coat of pigmented scales (e.g., Figs. 113 View FIGURES 108–113 , 384 View FIGURES 373–385 , 402 View FIGURES 402–406 ). Numerous egg striae with impunctate wide costae and narrow sulci lacking macroscopic punctures 3 but with a line of micropunctures on each side.................................................................................................. N. excisa and N. sjostedti operational complex, p. 130
1' S8 without structural modifications but several species have on S8 a pale anchor-shaped pattern between pigmented lightly sclerotised areas (e.g., Figs 123 View FIGURES 123–126 , 301 View FIGURES 294–304 , 345 View FIGURES 344–350 ). SSt and eggs variable............................................... 2
2 SSt not coiled or spiral, variable, usually pale and not densely coated with scales or spicules.......................... 3
2' SSt spiral or coiled, length varies from a partial circle (e.g., Figs. 167 View FIGURES 161–168 , 226 View FIGURES 223–226 ) to 1–6 complete rings (e.g., Fig. 126 View FIGURES 123–126 ). Inside with dense coat of pigmented scales or spicules, except on a narrow bare seam along concave edge. Next to the vagina the SSt may have a (probably extensible) section with folded soft cuticle and only a narrow band of scales on convex side, the scaly band widens distally until it occupies the entire width of the SSt (e.g., Fig. 99 View FIGURES 96–101 ). Eggs vary............................... 10
3 SSt comprising sections directed in opposite directions; base directed cephalad from origin on vagina, distal part pointing backward (e.g., Figs. 46 View FIGURES 42–49 , 50 View FIGURES 50–55 ). Eggs with a low number (<20) of ridges, entire surface of chorion sculptured in a similar way.. ................................................................................................... 4
3' SSt directed caudad from the origin on the vagina, not subdivided. Eggs variable................................... 5
4 Large and pale, with clear almost 20 mm long wings. S8 with short transverse subgenital plate. A narrow tube runs cephalad from the vagina, then curves back and turns into a flat slender leaf-shaped section which narrows caudally. Egg stout, with few straight ridges, operculum a sharply delimited conical cap. Stem of anchor solid but several filaments are distally attached ( Figs. 50–55 View FIGURES 50–55 )......................................................................... 10 N. View in CoL luluana Navás
4' Size variable, no subgenital plate. Vagina with wide dorsal opening into the flat bag-shaped SSt. Egg with 6–16 ridges, anchor mushroom-shaped, a cap on a single stem........................................... N. transvaalensis -group, p. 8
5 SSt is a short curved wide semi-tube, armature delicate, pale. Egg subspherical, not striate, with pale eclosion line, without collar ( Figs. 74–75 View FIGURES 69–75 )..................................................................... 15 N. coffea n. sp.
5' SSt and eggs different, variable.......................................................................... 6
6 SSt a semicircular tube which is much longer than the vagina and has only a narrow seam of scales along the convex periphery. Egg a slender ovoid with numerous punctate flat striae, operculum conical ( Figs. 66–68 View FIGURES 63–68 )............ 13 N. angolana n. sp.
6' SSt different and not significantly longer than the vagina, egg different........................................... 7
7 SSt is a curved funnel densely coated with scales. Origin on vagina very wide, the free end narrows gradually. Egg with low conical cap and approximately 12 straight ridges on the punctate chorion, only the narrow collar is not puncate ( Figs. 94–95 View FIGURES 91–95 )................................................................................ 17 N. planidorsum n. sp.
7' SSt and egg different.................................................................................. 8
8 SSt is a plump curved bag, inside it are numerous microtrichia. Egg ovoid, collar sessile and wide, ~ 20 straight costae and irregularly punctate sulci, the conical operculum with punctate grooves ( Figs 69–73 View FIGURES 69–75 )................. 14 N. pickeri n. sp.
8' SSt slender and almost straight, resembling a finger.......................................................... 9
9 Large-bodied, wings approximately 24 mm long. Egg drum-shaped, with low, sharply delimited cap, sides with approximately 16–18 flat, complexly structured dextrogyrous costae separated by narrow fissures. Collar integrated into contour of egg ( Figs. 59–62 View FIGURES 59–62 )........................................................................... 12 N. pallidogigas n. sp.
9' Smaller bodied, wings < 15 mm long. Egg ovoid, operculum rounded, 18 straight striae and sulci, the projecting collar with constricted base ( Figs. 81–82 View FIGURES 76–82 )........................................................ 16 N. duodeviginti n. sp.
10 SSt forming 1–2 rings, inside with needle-like microtrichia ( Fig. 109 View FIGURES 108–113 ). Egg slender, spindle-shaped, both poles narrow, no collar, with ~ 20 straight ridges ( Figs. 83–90 View FIGURES83–90 )..................... 17 N. View in CoL didita (Enderlein) and 18 N. juxtadidita n. sp.
10‘ Inside of SSt with dense coat of flat scales ( Fig. 113 View FIGURES 108–113 ), length varies from a short regularly curved fraction of a ring to long spirals of up to 5 superimposed rings, or very long narrow tubes forming a tangle ( Figs 126 View FIGURES 123–126 , 167 View FIGURES 161–168 , 335 View FIGURES 334–339 )................. 11
11 The entire egg surface is punctate (e.g., Fig. 112 View FIGURES 108–113 ), with 6–16 straight ridges (e.g., Figs. 100–101 View FIGURES 96–101 ). Some species with coarse punctation have ornaments on top of ridges (e.g., Fig. 111 View FIGURES 108–113 )................................. N. africana -group, p. 50
11' Impunctate costae or smooth ridges present on the chorion.................................................... 12
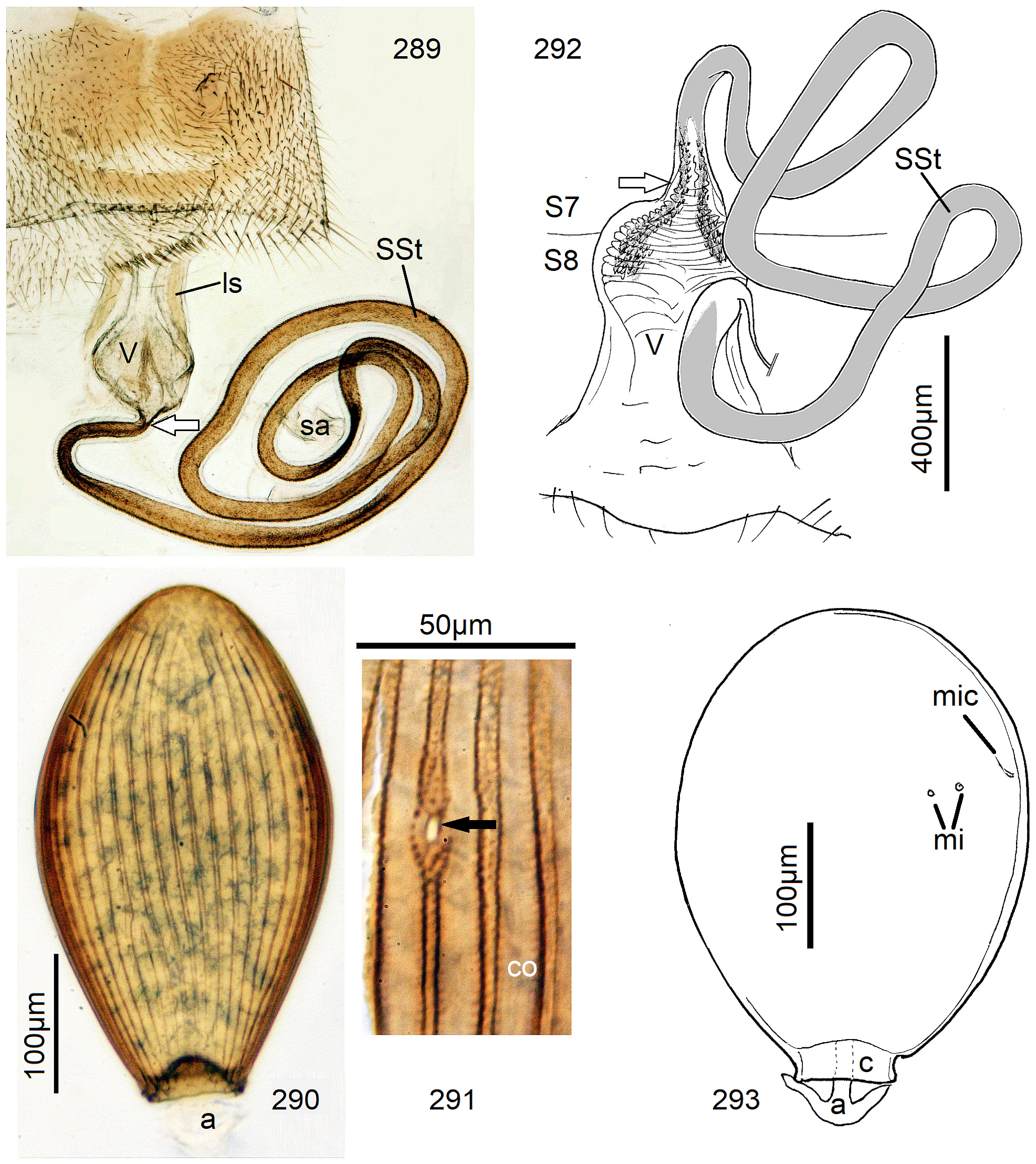
12 Egg striae are composed of impunctate raised costae and low-lying or incised punctate sulci (e.g., Figs. 110 View FIGURES 108–113 , 214 View FIGURES 213–214 , 291 View FIGURES 289–293 , 401 View FIGURES 397–401 ). Anchor mushroom shaped, a solid stem supports the cap (e.g., Figs. 7 View FIGURES 6–10 , 31 View FIGURES 26–32 , 392)................................... 13
12' Egg striation is caused by only raised ridges of variable height ( Figs. 303 View FIGURES 294–304 , 312 View FIGURES 309–312 ). Anchor resembling a mushroom but the cap rests on several thin delicate threads ( Figs 302 View FIGURES 294–304 , 308 View FIGURES 305–308 ), no solid stem present................ N. orthonema -complex, p. 116
13 Costae and sulci are of similar width, sulci with unordered punctation and freely visible micropyles (e.g., Fig. 257 View FIGURES 250–257 ), except in species where costae bear crest of flanges ( Figs. 324–325 View FIGURES 323–331 )................ N. spio - and N. dubia -complexes, p. 87, p. 130
13' Costae are 2 or more times wider than the sulci which lack macroscopic punctures but have a row of micropunctures on each side (e.g., Figs. 343 View FIGURES 340–343 , 350 View FIGURES 344–350 , 360 View FIGURES 351–361 ). Micropyles are not exposed, in some species they are completely concealed............ 14
14 S8 with a pale anchor-shaped mark, egg without collar....................................................... 15
14' S8 without pattern, caudally not modified. Egg shape variable, with or without collar, striae straight or levogyrous (anticlockwise, spirals in side view running from top left to bottom right). In mounted eggs regular rows of micropunctures are visible at magnifications ≥ 200X. Micropyles variable............... N. excisa and N. sjostedti -operational complex, part, p. 130
15 S8 caudally not prolonged, SSt short, approximately twice as long as vagina. Egg with many straight striae and a pale ring at base of operculum ( Figs. 344–350 View FIGURES 344–350 )........................................................ 61 N. lineata View in CoL n. sp.
15' S8 caudally extended, distal part of anchor-shaped mark parabolic............................................. 16
16 Egg drum-shaped, operculum wide and smooth, sides of egg with line-like fissures forming dextrogyrous spirals (clockwise, in side view running from top right to bottom left) ( Figs. 357–361 View FIGURES 351–361 )........................... 62 N. conradti View in CoL (Enderlein)
16' Egg ovoid, striae are levogyrous spirals, the narrow operculum with shallow cells ( Figs. 362–364 View FIGURES 362–364 )..... 63 N. muyukae n. sp.
Key to females with non-striate eggs
1 S8 caudally with a nail....................................................................... 2
1' No nail on female S8........................................................................ 3
2 SSt a long spiral. Nail longer than wide at base ( Figs. 369–371 View FIGURES 365–372 )..................... 64 N. excisa View in CoL Klapálek
2' SSt shorter, about 1 ring. Nail short and truncate (Figs. 391–393)...................... 68 N. simplex n. sp.
3 SSt short and wide, a semi-tube with delicate scales. Egg drop-shaped, with a visible eclosion line ( Figs. 74–75 View FIGURES 69–75 )............................................................................. 15 N. coffea n. sp.
3' SSt a curved or coiled flat band, length varies from a fragment of a circle to a long spiral. Inside with dense coat of brown scales.
Eggs variable, no visible eclosion line.......................................................... 4
4 Egg without trace of a collar. Egg pear-shaped, diameter almost doubles near midlength ( Figs. 158–160 View FIGURES 154–160 ).................................................................................. 28 N. pirus n. sp.
4' Anchor pole of egg surrounded by a sclerite ring (e.g., Figs 156–157 View FIGURES 154–160 ) or with a proper collar. Eggs subspherical or of regular ovoid shape....................................................................... 5
5 Base of vagina with transverse folds (tf), SSt forming approximately 1 ring (e.g., Figs. 181, 186, 188 View FIGURES 181–192 )............................................................................. N. pilulifera-complex , p. 70
5' No transverse folds in base of vagina........................................................... 6
6 SSt present as a very long coil ( Figs. 292–293 View FIGURES 289–293 ).................................. 51 N. sambarua n. sp.
6' SSt short, forming half of a circle.............................................................. 7
7 Egg stout, barely 1.5 times as long as wide, only near the poles with short radial ridges between irregular deep punctures ( Figs. 223–226 View FIGURES 223–226 ).................................................... 42 N. bipolaris n. sp.
7' Egg slender, with obsolete striae: costae narrow and very low, hardly raised above the sulci which have unordered punctation (Figs 227–230)..................................................... 43 N. schuelei n. sp.












No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Neoperla Needham, 1905
| Zwick, Peter & Zwick, Andreas 2023 |
Oodeia Klapálek, 1921: 321
| Klapalek, F. 1921: 321 |
Formosita Klapálek, 1914:118
| Klapalek, F. 1914: 118 |
Formosina Klapálek, 1913: 117
| Klapalek, F. 1913: 117 |
Tropidogynoplax
| Zwick, P. 1973: 499 |
| Enderlein, G. 1910: 141 |
Ochthopetina
| Zwick, P. 1973: 496 |
| Enderlein, G. 1909: 324 |
Javanita Klapálek, 1909: 224
| Klapalek, F. 1909: 224 |
Neoperla
| Needham, J. G. 1905: 108 |
Pseudoperla
| Pictet, F. J. de la & Hagen, H. A. 1856: 37 |