
Limnosida frontosa Sars, 1862
|
publication ID |
https://doi.org/10.11646/zootaxa.3764.1.4 |
|
publication LSID |
lsid:zoobank.org:pub:E1CA4935-5A1D-4BDD-9847-830FE4570369 |
|
DOI |
https://doi.org/10.5281/zenodo.5658709 |
|
persistent identifier |
https://treatment.plazi.org/id/E1567004-FFC7-FF8A-F198-15BBFE7FFC3E |
|
treatment provided by |
Plazi |
|
scientific name |
Limnosida frontosa Sars, 1862 |
| status |
|
Limnosida frontosa Sars, 1862 View in CoL
( Figs. 1–7 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 )
Sars, 1861 (1993): 18–20, Pl. 6, 1–5.
Sars, 1862: 259–261;
Sars, 1865: 37–41, Tab. II, fig. 1–15;
Richard, 1895: 341–343, Pl. 15, fig. 6, 10, 17; Lilljeborg, 1901: 29–34, Tab. II, fig. 4–11, Tab. III, fig. 1–5; Rylov, 1935: 105, Taf. 14, Fig. 135a, b; Šramek-Hušek, 1962: 182–184, Obr. 63; Manuilova, 1964: 108, Fig. 26, 1–7;
Flössner, 2000: 39–41, Abb. 15a–i;
Korovchinsky, 1992: 5, Figs. 18–25;
Korovchinsky, 2004: 256, Fig. 81, 1–7.
Body measurements of specimens of seven populations are presented in Table 1.
Description. Parthenogenetic female. Body oval-elongated with comparatively large head (length 34.6–45.3 % and height 29.4–40.7 % of body length) of peculiar triangular shape ( Fig. 1 View FIGURE 1 A–C), narrowing frontally and ventrally, and with no rostrum. Large eye (7.7–12.7 % of body length), situated in the narrowed ventral part of head. Small black ocellus near the eye.
Antennules small ( Fig. 1 View FIGURE 1 D) with 9 aesthetascs with constrictions and short sensory seta in apical position, situated in two groups: four aesthetascs and sensory seta on the anterior small apical prominence of the antennular base and five aesthetascs positioned posterior to them.
Swimming antennae long (61.4–80.6 % of body length) with massive basipodite (40–46 % of body length), having long setulated seta on dorsal side of its base ( Fig. 1 View FIGURE 1 G). Distal end of antennal basipodite bears stout spine on its outer middle side, small bifurcated prominence dorsally in front of upper branch, and small dorsal spine-like seta better seen from the inner side ( Fig. 1 View FIGURE 1 E, F, L). The upper three-segmented antennal branch (exopodite) is longer (29–32 % of body length) with segments of different size, the proximal of which is smallest (5 % of body length) and lacks setae while the following two are longer (10 and 16 % of body length) and setulated with three (6.7, 10.0 and 13.6 % of body length) and seven (14.2, 14.2, 14.2, 13.6, 13.0, 11.5, 11.5 % of body length from proximal to distal) long, two-segmented setae, respectively. Two latter segments bear apically one small spine each ( Fig. 1 View FIGURE 1 H, I; 4B). In lower shorter (19–22 % of body length) three-segmented antennal branch (endopodite), proximal and distal segments are small while second one is most developed (2.5, 13 and 4.5 % of body length) bearing apically one long seta (13.3 % of body length) and strong spine ( Fig. 1 View FIGURE 1 J). Distal segment of this branch bears four setae (14.2, 14.2, 12.4, 13.6 % of body length, counting from ventral to distal side) and one spine ( Fig. 1 View FIGURE 1 J, K). Proximal segments of both branches clearly articulate with neighboring ones externally while internally they seem completely fused with the latter ( Fig. 1 View FIGURE 1 L). Formula of antennal setae: 0–3–7 / 0–1–4. All setae of swimming antennae armed uniformly with stout setules of swimming type.
Mouth parts consist of two maxillules (mx I), each with 9–11 densely situated feathered setae incurved forward, two small maxillae (mx II) situated just near base of tl I ( Fig. 2 View FIGURE 2 F) and having short setulated subapical seta, and two mandibles. Molar surface of right mandible ( Fig. 4 View FIGURE 4 D) is oval-elongated, with a group of one large thorn and few smaller ones near it (pegs—P) posteriorly. A row of some comb-like prominences (pales—Pa) goes along dorsal margin while a row of sharpened prominences (fishbones—Fe) and one large rounded outgrowth are situated along ventral margin. Central part of molar surface with 10–11 diagonals (D) supplied with numerous pores. Left mandible ( Fig. 4 View FIGURE 4 E) seems somewhat wider, with a group of pegs, consisting of two large and some small ones near them, at its dorso-posterior angle. Pales form a practically fused row, lowering anteriorly. A group of two-three large fishbones near pegs, the outer of which is clearly branched while inner one(s) looks like distally widened plate with comb-shaped margin. Another prominent group of fishbones with comb-shaped apical ends inclined anteriorly is situated on large outgrowth between pegs and pales. Diagonals, also with pores, are poorly delineated.
Shell with more or less arched dorsal side, connecting with rather high posterior valve margin through emarginated dorso-posterior angle. More or less prominent longitudinal fold separates upper and lower parts of shell ( Fig. 1 View FIGURE 1 A, B). Posterior valve margin with prominent emargination in its middle part, the form ( Fig. 1 View FIGURE 1 M) and depth (1.5–5.2 % of body length) of which vary among specimens. Row of tiny spinules runs marginally and submarginally along its lower part. Ventral valve margin is almost straight, with numerous small uniform denticles along it. Comparatively large denticle, naked or spinulated, is situated on ventro-posterior corner of both valves ( Fig. 2 View FIGURE 2 B–E).
Six pairs of thoracic limbs, all with comparatively small oval epipodites. Their structure and armament are shown in Table 2 View TABLE 2 . Exopodites of tl I–tl V are widened terminally and show signs of segmentation ( Fig. 5 View FIGURE 5 A, C). Endopodites (this is a conditional term according to Smirnov and Kotov (2010)) of these limbs are more or less clearly subdivided in four segments, the proximal of which is largest, bearing numerous filtering setae, while three terminal segments bear two-four such setae. Terminal segment from tl I to tl V and subterminal segment from tl II to tl V bear also outer setae each, similar to those of exopodite, of which the former one is especially long ( Fig. 5 View FIGURE 5 A, B, D, indicated by arrows). Large proximal segment of endopodite of tl I also possesses terminally one short naked seta ( Fig. 5 View FIGURE 5 A: “ns”). A row of comparatively long rough setules along inner side of endopodite and a group of spines on inner anterior basal side of tl I ( Fig. 5 View FIGURE 5 A, C). Gnathobase of tl I ( Fig. 5 View FIGURE 5 A, B) has outer row of six-eight finely setulated filtering setae; inner long, two-segmented setae (I) on distal corner with setules along its length, small spine-like seta near its base, and another smaller curved setulated seta (c), between these there are two soft, sensilla-like prominences ( 2i) of different sizes ( Fig. 5 View FIGURE 5 B). Two groups of long spinules situated just proximal to them ( Fig. 5 View FIGURE 5 A). Gnathobases of tl II to tl V ( Fig. 5 View FIGURE 5 C) with row of 14–23 finely setulated filtering setae, ending proximally with one naked seta (p), and distally with two setae of different types, one of which is long and twosegmented (I), resembling that one of tl I but differing in presence of stout spinules distally, and the other one strongly modified, being hooked terminally and denticulated laterally (J) ( Fig. 5 View FIGURE 5 C, E). Tl VI ( Fig. 5 View FIGURE 5 F) is small and strongly modified. Its exopodite ( Fig. 5 View FIGURE 5 G) has been reduced to a terminal plate, armed with five terminal and one lateral setae and some long spinules. Endopodite is almost entirely fused with exopodite, bearing seven similar setae with long setules and one thorn between the proximal two and distal five. Gnathobase with two long setae and three thorns of differing shapes. Surface of endopodite and gnathobase with rows of long setules.
Limb pairs Exopodite (apical + Endopodite Gnathobase Epipodite
lateral setae)
I 7 + 4 ( n 3+1)+( n 3)+( n 3)+( n 18−21) ( n 6−8)+(b+I+ 2i +c) + Postabdomen comparatively massive, short (12.2–17.1 % of body length), tapering distally, high (height in its proximal part 8.3–12.2 % of body length) ( Fig. 2 View FIGURE 2 A) with long two-segmented postabdominal setae (58.7–63.6 % of body length) sitting on two separate bases. Their longer distal parts bear very thin and sparsely situated setules of “sensory” type. Small triangular prominence situated anterior to the setae on the dorsum of abdomen. Lateral and dorsal sides of postabdomen with groups of small spinules ( Fig. 4 View FIGURE 4 C). Postabdominal claws rather long (9.7–16.4 % of body length) and massive with 7–16 long spines, diminishing in size proximally and varying considerably in size and shape ( Figs. 2 View FIGURE 2 H–R, 3A–X, Z). Claw outer lateral sides with long rows of spinules of smaller or larger size ( Fig. 4 View FIGURE 4 F, G) while on dorsal and inner sides these are less numerous ( Fig. 4 View FIGURE 4 H).
Juvenile females. They have small body size ( 0.56–0.90 mm), fewer antennal setae (0–2–7 / 0–1–4), which sometimes are accompanied by rudiment of proximalmost seta on second segment of upper antennal branch (larger juveniles often have full number of antennal setae), and fewer number of spines on postabdominal claws (5–8) ( Fig. 3 View FIGURE 3 Y).
Gamogenetic female. They are similar to parthenogenetic females with respect to all diagnostic features, differing from them only in presence of dark widely-oval resting eggs ((0.21–0.26) x 0.18 mm) in brood pouch ( Fig. 2 View FIGURE 2 G).
Male. General body shape ( Fig. 6 View FIGURE 6 A) is subtler than in females with low dorsal side of shell, having prominent dorso-posterior angle. Head with rounded anterior part and slightly larger eye (8.7–13.2 % of body length). It bears very long (57.3–82.2 % of body length), more or less S-curved antennules with massive proximal part ( Fig. 6 View FIGURE 6 B, C), bearing two outgrowths and nine aesthetascs situated on a small prominence on inner side of antennules in two groups of four and five ( Fig. 6 View FIGURE 6 D). Proximal outgrowth large, cone-shaped with slightly widened apical end directed backwards; distal one, smaller and usually rounded, quite variable in size and shape ( Fig. 7 View FIGURE 7 A–F, H–V), occupies anterior semiinternal position. A transparent stick-like solid structure ( Sars 1865: Tab II, 14, 14a), which probably fulfils a supportive function, lies inside the basal parts of the antennules. Distal portion of antennules long, narrow, tapering to cone-shaped or finely-pointed terminus, and covered with numerous densely situated fine setules ( Figs. 4 View FIGURE 4 A, 6E, F). Swimming antennae (64.3–75.0 % of body length) identical to those of female ( Fig. 6 View FIGURE 6 I). First thoracic limbs with large clasping organ ( Fig. 6 View FIGURE 6 J–L), consisting of a large, probably movable, hook, a group of tiny denticles adjacent to it, and a prominent thorn located among them external to the hook ( Fig. 6 View FIGURE 6 L). Three filtering setae and one massive two-segmented naked seta are attached to small outgrowth situated internally near clasping hook ( Fig. 6 View FIGURE 6 J, K). All other setae as in female, including small naked seta (“ns”) near distal end of proximal row of filtering setae.
Juvenile males. In smaller males, antennules somewhat like that of females ( Fig. 6 View FIGURE 6 G) but the sensory seta seems thicker and stays separate from aesthetascs. In larger males, antennule base enlarges, stretches and fuses with modified and enlarged sensory seta ( Fig. 6 View FIGURE 6 H).
Size. Parthenogenetic female body length: 1.0– 2.03 mm; up to 2.40 mm according to Sokolova (2005). Male body length: 1.0– 1.87 mm.
Individual and interpopulation variability. In adult females, measured structures usually showed moderate variability among individuals, but variablility of diameter of eye, height of postabdomen, length of postabdominal claws, and especially depth of embayment of posterior valve margin was high (see Table 1). Number of filtering setae of endopodite and gnathobase of thoracic limbs varied as well (see Table 2 View TABLE 2 ). Differences between mean values (M) of features, both significant (p <0.001; p <0.01) and insignificant, of specimens from different regions showed random distribution within the range of L. frontosa .
Spines of postabdominal claws, especially distal ones, differed in size and degree of inclination to claw. Specimens of some populations had comparatively short and weakly inclined spines ( Fig. 2 View FIGURE 2 H–J, Fig. 3 View FIGURE 3 O, R), while others had both less inclined and more inclined spines ( Fig. 2 View FIGURE 2 K, L, N, O), or had only strongly inclined spines ( Fig. 3 View FIGURE 3 A–G, I–M, Q, U, V, X, Z).
While the proximal outgrowth of male antennules demonstrated morphological stability, the distal one varied considerably in size and shape ( Fig. 7 View FIGURE 7 A–F, H–V). Conspicuous intra- and interpopulation variability of this feature was frequently observed ( Fig. 7 View FIGURE 7 C–E, N–O, P–Q, R–S); this was especially prominent in specimens from Kunovatsky sor (Lower Ob’ River basin, Western Siberia).
In some specimens, malformed postabdominal claws, spines, and antennules ( Figs. 3 View FIGURE 3 S, T, W; 7F, G) were observed.
Remarks. Sars (1861 (1993), 1862) found and originally described L. frontosa only from Lake Koldbjørnsvigsjønen in south-eastern Norway. The type series ( syntypes) is stored in ZMUO (M. Christiansen, pers. comm.) and was not accessible for this study. Norwegian specimens used in this study, initially studied and labeled by G.O. Sars, originate from another Norwegian location, Lake Stensrudvann.
The original description of L. frontosa was brief and incomplete, lacking figures and including only females. (Although figures were present in the unpublished manuscript ( Sars, 1861 (1993)), some of them were later lost). In addition, comparison of the present data with those by Sars revealed some inaccuracies. Initially, he counted eight antennular aesthetascs in two groups of four, and later, in his more detailed description ( Sars 1865), seven aesthetascs were incorrectly noted, three in the anterior group and four in the posterior one. Thoracic limbs were poorly described and sketchily drawn: in the female only tl III and tl VI and in the male only tl I.
Region and number Spines of postabdominal claws Distal outgrowth of male’s antennules of locality
short and weakly inclined long and strongly inclined short long NW 1 +
NW 2 +
NW 3 +
NW 4 +
NW 5 + + NW 6 +
NW 7 + + NW 8 + + NW 10 +
VR 11 + + + VR 12 +
VR 14 + + NE 15 + + + NE 16 + +
NE 17 + + + WS 18 +
WS 20 + + WS 21 + + WS 22 + + ES 23 + + ES 24 + + + ES 25 +
ES 26 +
ES 27 + + + ES 29 + + ES 30 + + FE 33 +
FE 34 +
FE 35 +
FE 37 +
FE 38 +
FE 41 + + FE 42 + + FE 43 + + FE 44 +
FE 46 + + FE 47 +
FE 48 +
Lilljeborg (1901) was even less detailed than Sars and almost fully repeated his mistakes. He described “7−8 aesthetascs” of the antennules and presented sketches only of selected thoracic limbs (tl I and tl VI in female and tl I in male). Shortly thereafter, Scourfield (1905) first described the antennules of Limnosida correctly.
Some variable morphological features described above, i.e. spines of postabdominal claws, distal outgrowth of male’s antennules, denticle on postero-ventral angle of shell valves, and emargination of posterior shell margin, seemed likely to have potential taxonomic significance. However, the latter three features proved in general to be randomly distributed over the different regions of the continent which is also characteristic for all meristic features presented in Table 1. Although from Table 3, data suggest that males with a long distal outgrowth of the antennules are absent from the Far East, too few males were collected from this region to make this a certainty.
On the other hand, distribution of types of spinulation of the postabdominal claws shows, in general, a more regular trend (see Table 3). Specimens with comparatively short and weakly inclined distal spines (“north-western type ”) are distributed mostly in the north-west of Eurasia (Fennoscandia, including the north-west of Russia) (populations 1–10 in “Material and Methods”) ( Fig. 2 View FIGURE 2 H–J, P), while specimens with comparatively long and strongly inclined spines (“eastern type ”) dominate in Eastern Siberia (Yakutia) (populations 23–30) and the Far East (populations 33–48) ( Fig. 3 View FIGURE 3 D, E, J–Q, T–V, X, Z) as well as in the north of Western Siberia (populations 18– 22) ( Fig. 3 View FIGURE 3 A, C). Further, similar specimens were encountered from the middle part of the Volga River basin (populations 11–14) ( Fig. 3 View FIGURE 3 I).
In addition, specimens from two populations (15 and 16) from the north-east of European Russia (Komi Republic) ( Fig. 2 View FIGURE 2 K, N) and from one population from Western Siberia (17—Kunovatskiy sor) ( Figs. 2 View FIGURE 2 M, 3B, C) demonstrated presence of claw spines of both types. Also, specimens from two localities from the Lower Amur River (Far East—41, 42) ( Fig. 3 View FIGURE 3 O, R) have the claw spines of “north-western type ” and specimens from the Pregel River (Kaliningrad area, Russia) possess long and rather inclined spines ( Fig. 2 View FIGURE 2 R) of “eastern type ” (but here only two juveniles were present).
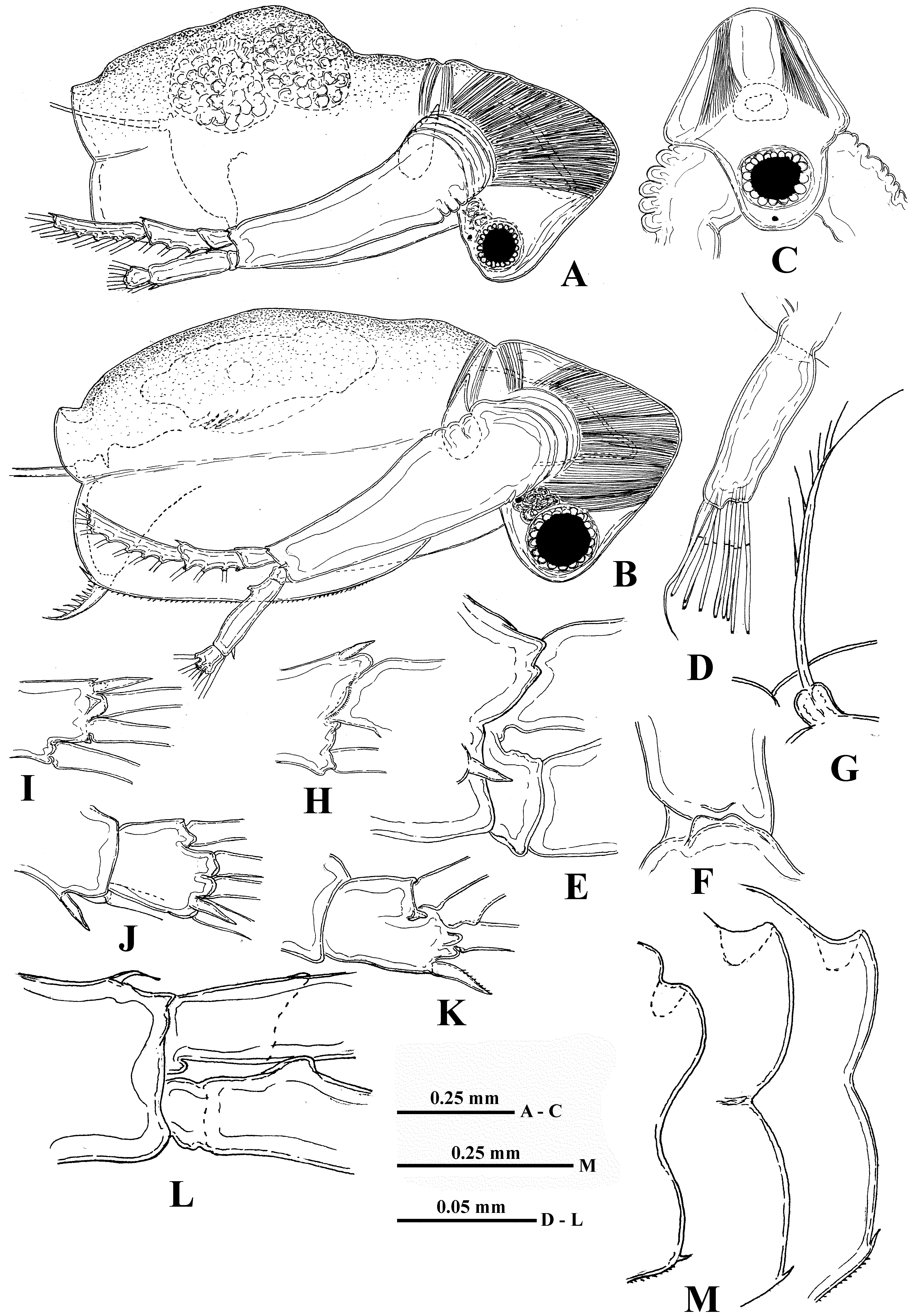
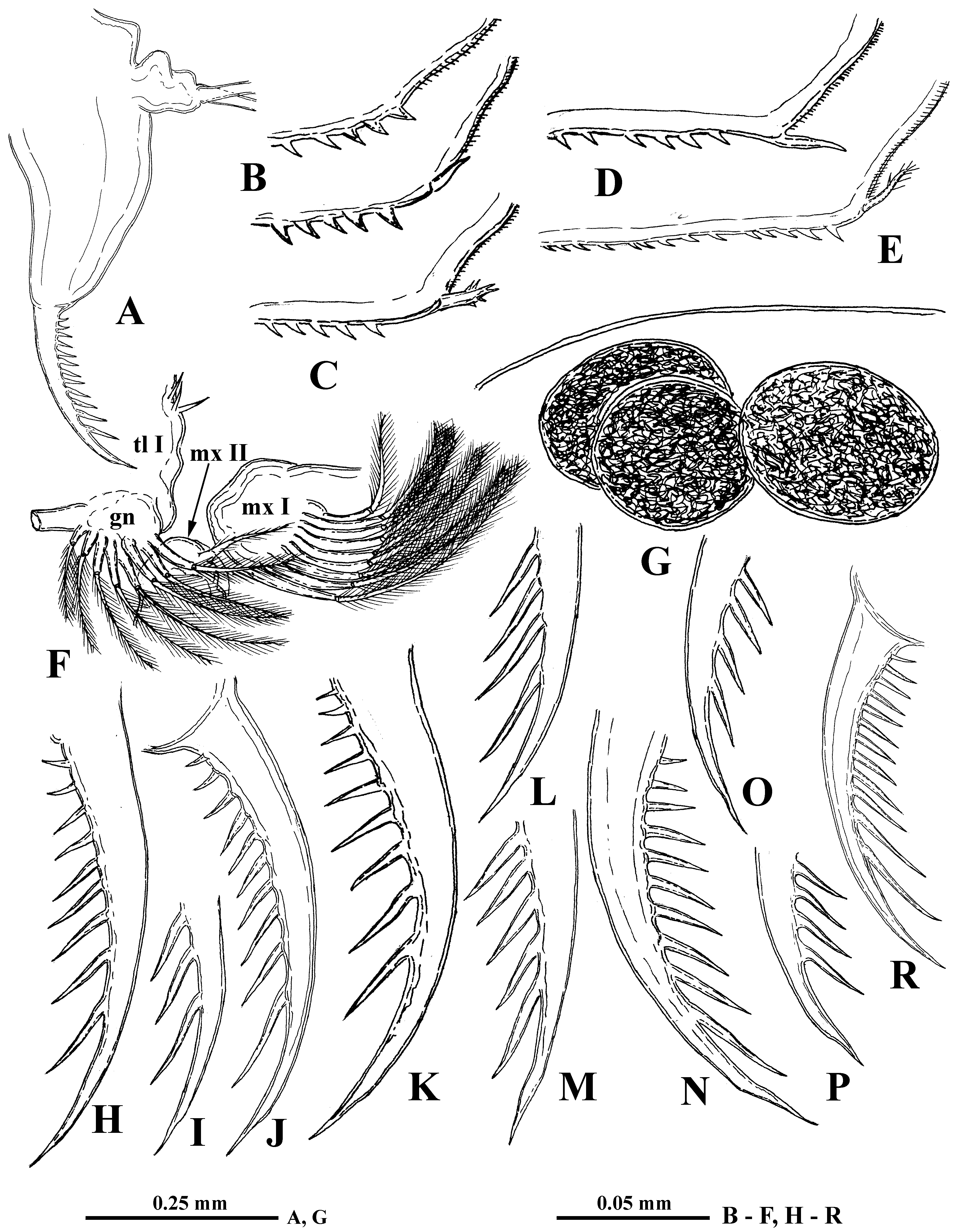

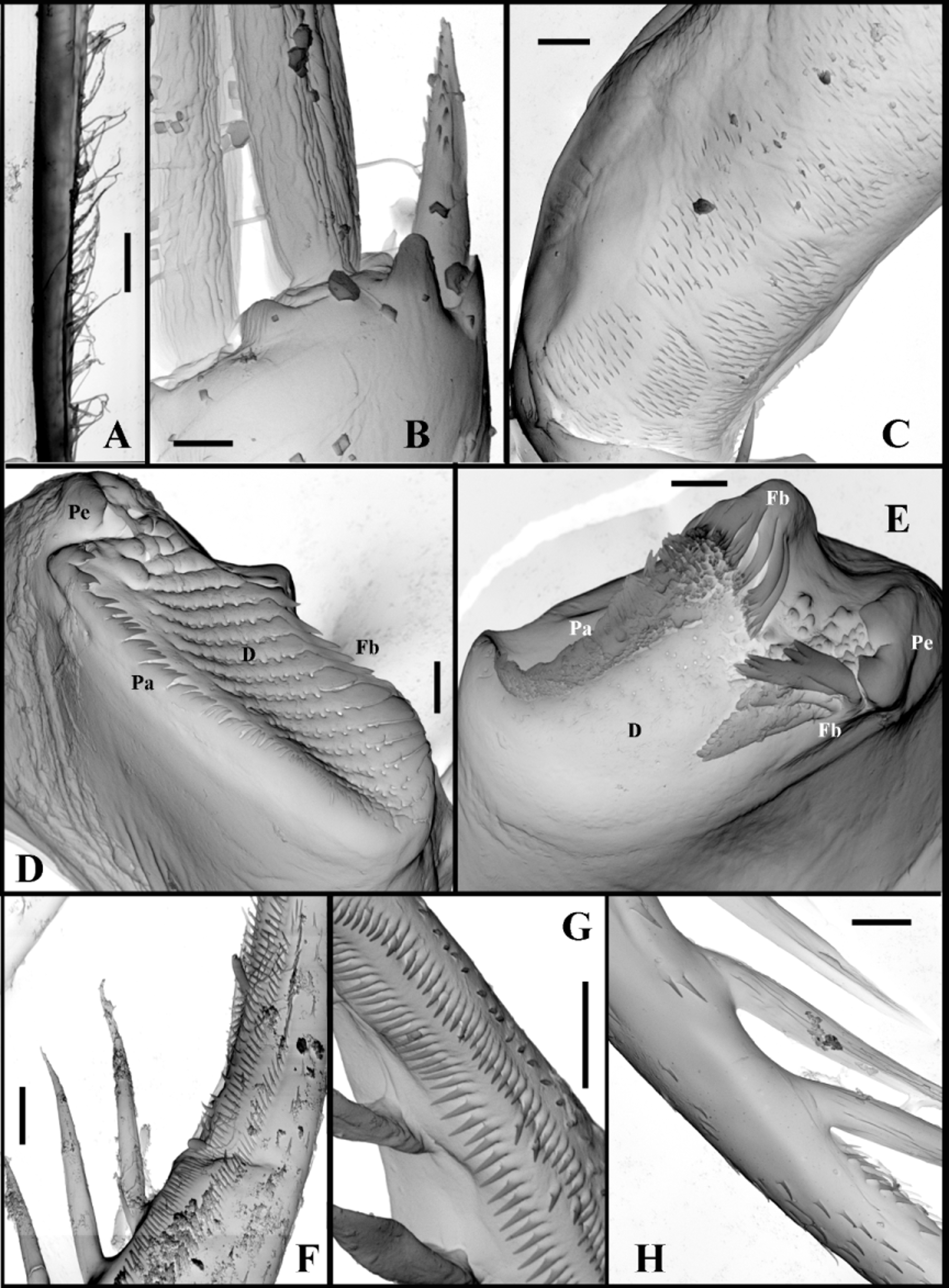
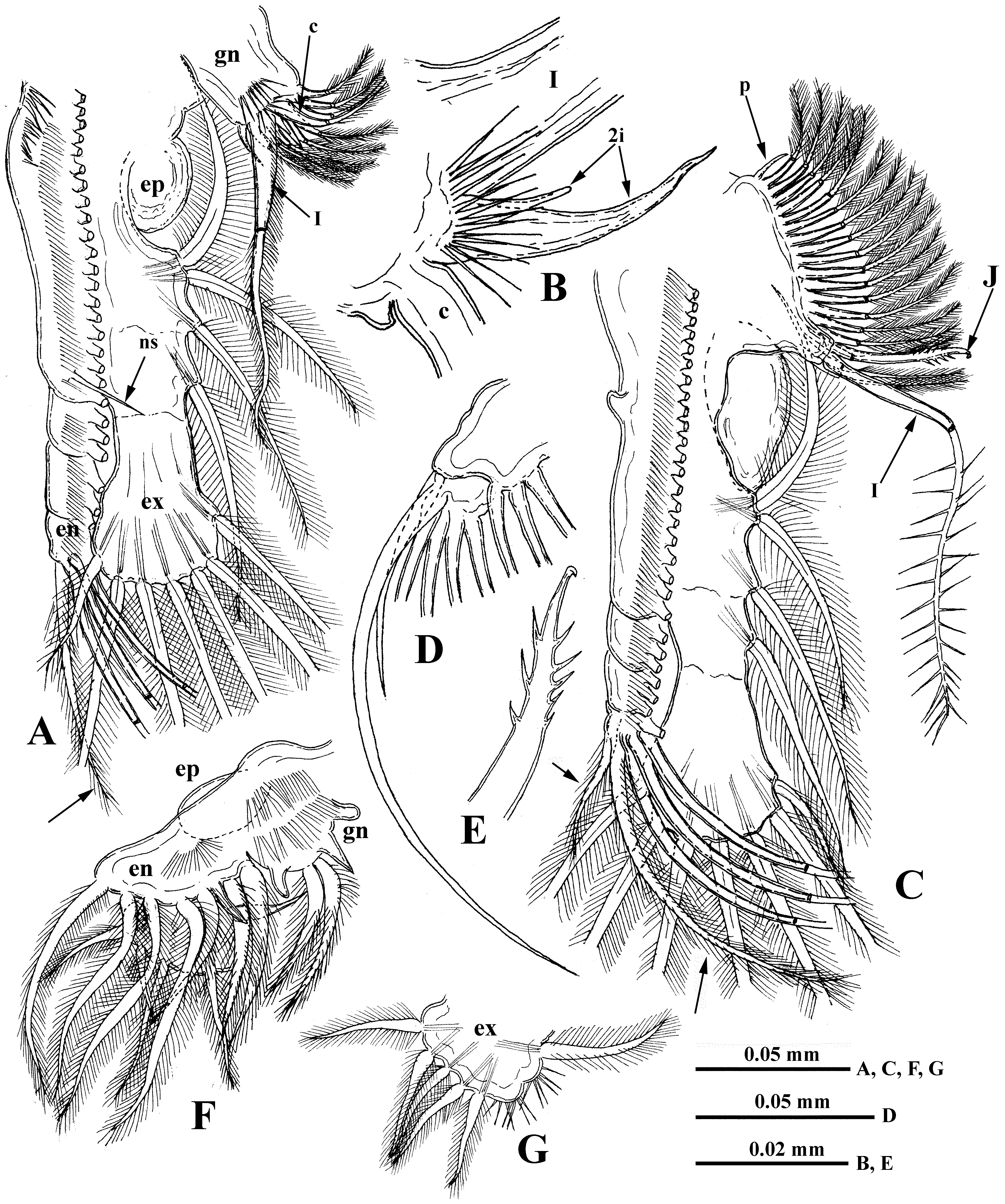
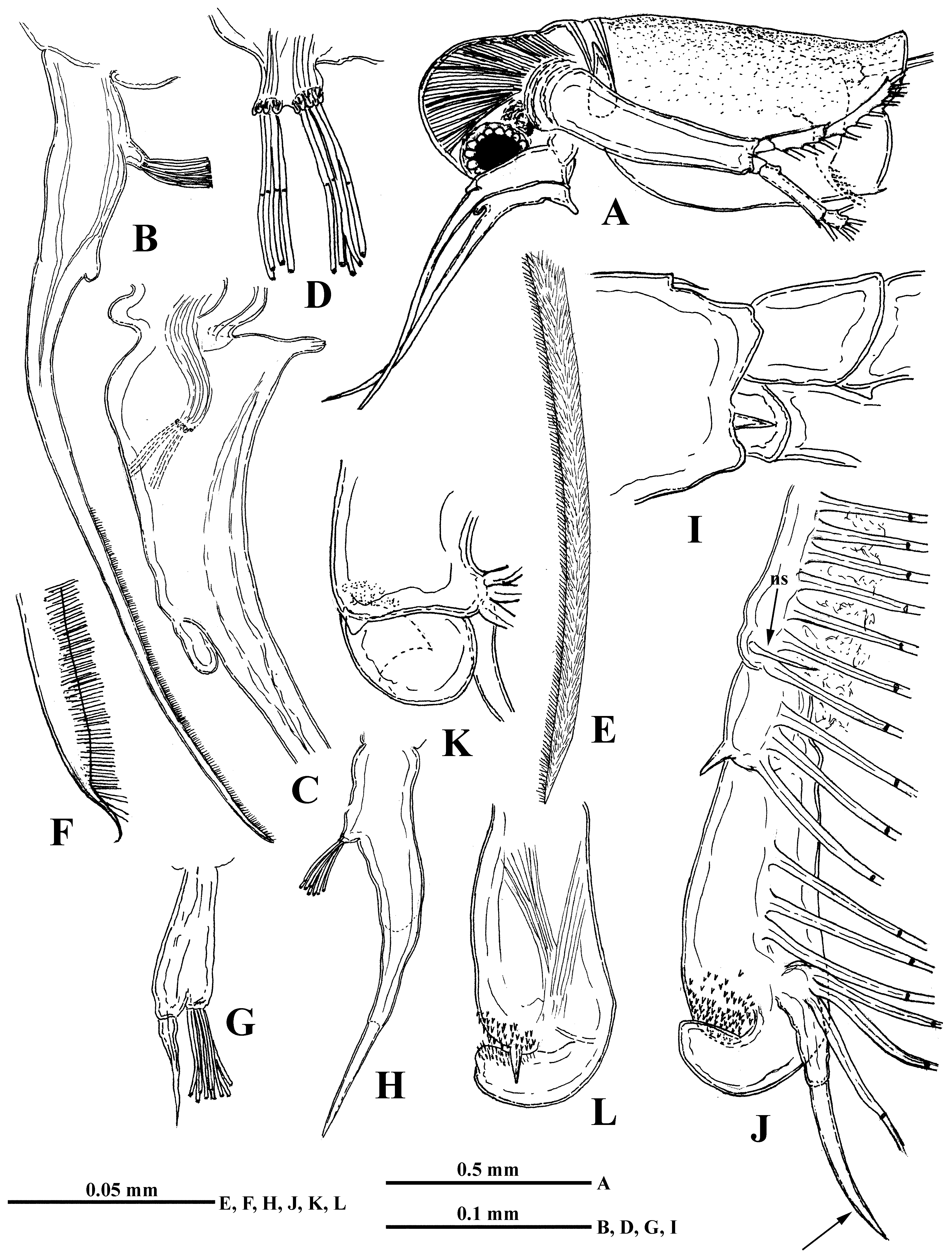
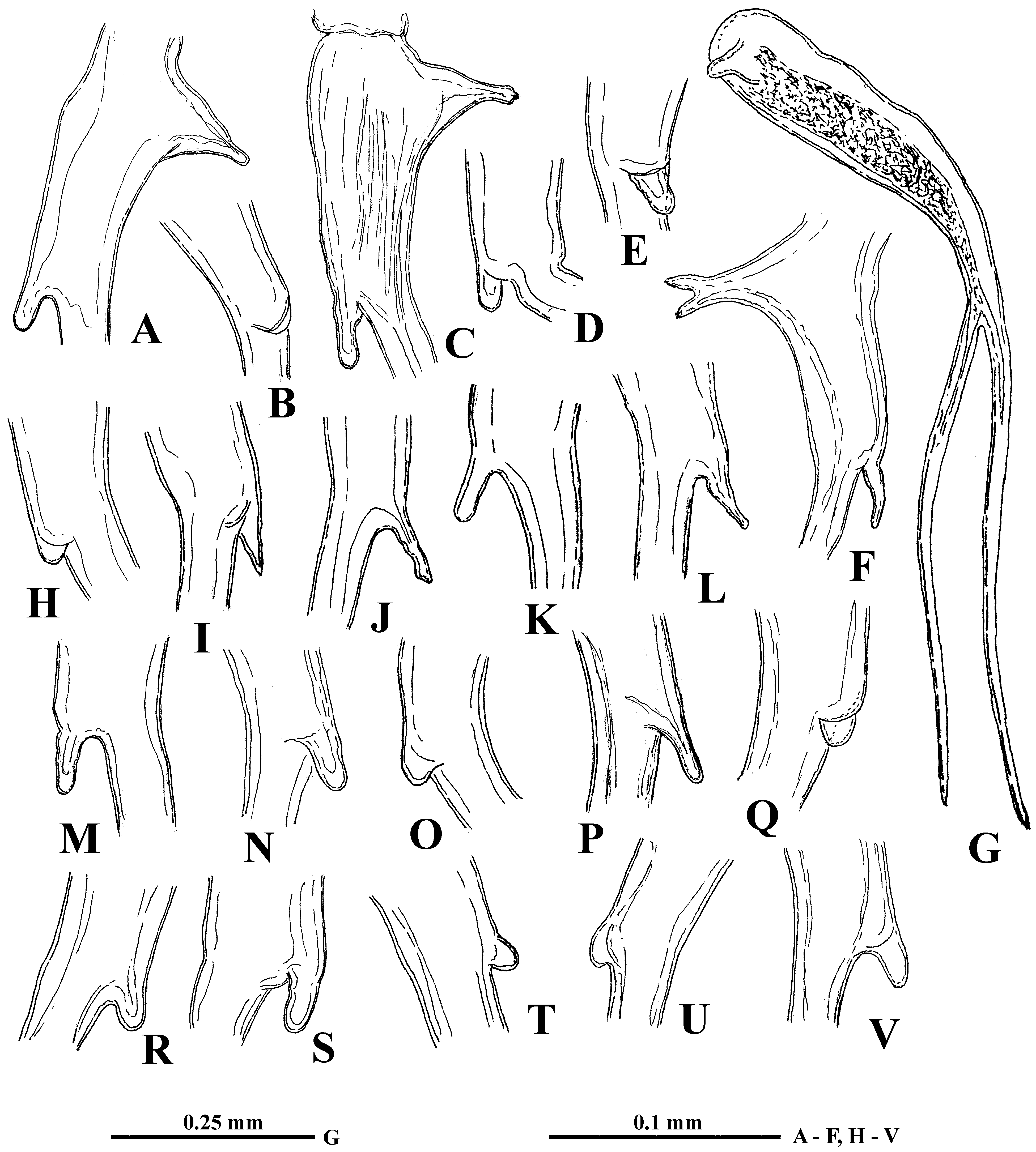
TABLE 2. Data on structure and armament of thoracic limbs of Limnosida frontosa (specimens from the Neva River estuary (Finnish Gulf, August 8, 2006).
| II | 7 + 4 | ( n 4+1)+( n 3+1)+( n 3)+( n 18−21) ( n 17−23)+(I+J)+p | + |
|---|---|---|---|
| III | 7 + 4 | ( n 4+1)+( n 3+1)+( n 3)+( n 19–20) ( n 16)+(I+J)+p | + |
| IV | 7 + 4 | ( n 4+1)+( n 3+1)+( n 3)+( n 16) ( n 14)+(I+J)+p | + |
| V | 6 + 3 | ( n 3–4+1)+( n 2–3+1)+( n 14–15) ( n 14)+(I+J)+p | + |
| VI | 5 + 1 | 7 + one thorn 2 + three thorns | + |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |