
Brachyorrhos Kuhl
|
publication ID |
https://doi.org/10.5281/zenodo.5350190 |
|
persistent identifier |
https://treatment.plazi.org/id/E37587A2-7E70-FFD5-FC7D-E8F0FED158BA |
|
treatment provided by |
Tatiana |
|
scientific name |
Brachyorrhos Kuhl |
| status |
|
Brachyorrhos Kuhl in Schlegel, 1826
Coluber Linnaeus, 1758: 128 View Cited Treatment
Brachyorrhos Kuhl in Schlegel, 1826: 236
Brachyorrhus Agassiz, 1847 : Emendation of Brachyorrohs Calamaria (in part) Schlegel, 1837: 25
Brachyorros Doria, 1874: 329. Error for Bracyhorrhos Atractocephalus Sauvage, 1879: 62
Brachyura Edgar & Lilley, 1993: 136
Type species. — Coluber albus Linnaeus by original designation
Comment on some taxonomic confusion. — The names Brachyorrhos kuhlii Boie, 1827 , Coluber brachyurus Shaw, 1802 , and Calamaria brachyorrhos Schlegel, 1837 are in fact based on a specimen of Atractus trilineatus (see Hoogmoed, 1982), a Neotropical dipsadids, but have been considered synonyms of Brachyorrhos albus by various authors. Macleay (1877) described Mainophis robusta from Papua New Guinea and Boulenger (1893) placed this snake in the synonymy of Brachyorrhos albus . The type specimen is in fact the elapid Furina tristis (Shea & Sadlier, 1999) , hence this name should be disregarded. All of these names have appeared in recent synonymies of Brachyorrhos (Iskandar & Colijn, 2001; Murphy, 2007; The Reptile Database (online), by Uetz, 2012) in part because of the complex nature of the confusion and the relatively obscure places the information has been published. Additionally, Heij et al. (1997) has confused Brachyorrhos albus with Boiga irregularis , suggesting that it eats bird eggs. Andersson (1899) reported that the type specimen is as described by Linnaeus, except that it has 27 subcaudals instead of 20, and that the fifth upper labial is divided (presumably horizontally divided) as opposed to what Boulenger (1893) reported for other specimens.
Diagnosis. — Brachyorrhos species are the only homalopsid snakes known to have bilobed nasal scales separated by the rostral—a rostral that is as broad as tall—and a divided internasal behind and dorsal to the nasals. Members of this genus can be distinguished from Calamophis (with which it has been long confused) by the lack of dorsal scale row reduction ( Brachyorrhos has 19-19-19, Calamophis has 19-19-17); dorsal scales are ovate, slightly imbricate or juxtaposed compared to dorsal scales in Calamophis ; a longer snout, a divided nasal scale, and a tail that lacks a constriction at its base and tapers gradually to the tip.
Description. — Small to medium snakes (adult size 30–74 cm) with a pointed head indistinct from the cylindrical, muscular body, and a relatively short but tapering tail. The nasal scale is bi-lobed with the anterior lobe larger and containing the valvular nostril. Loreal usually fused with the prefrontal and preocular (a preocular is present in B. albus ). One pair of chin shields makes contact with the first four lower labials. Five to seven upper labials, third, fourth, or 3 + 4 in orbit. Six to eight lower labials, first pair makes medial contact, fourth is the longest, four contact first pair of chin shields. Smooth dorsal scales in 19 rows on the neck, at midbody, and just anterior to the vent, thus there is no posterior reduction in dorsal rows. Scales are ovate and juxtaposed or very slightly imbricate. Ventrals 159–195; divided subcaudals 18–42, anal scale divided. Males tend to have lower ventral counts, higher subcaudal counts, longer tails, and smaller SVLs than females (see Table 3 for morphometrics).
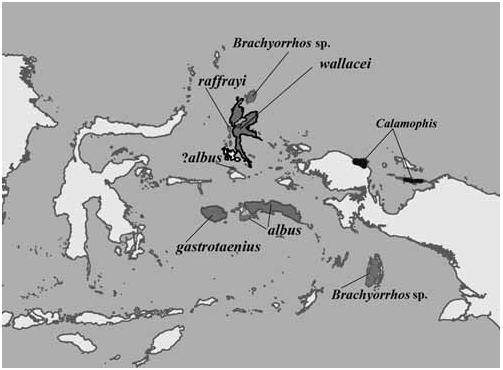
Distribution. — Literature reports of wide-ranging and often erroneous localities for Brachyorrhos have added confusion to the actual identity of these snakes. Despite the report of Brachyorrhos on Sumatra by Bleeker (1857a), Haas (1950) and David and Vogel (1996) found no evidence that it occurs there. Many of Bleeker’s records, including the Sumatra locality, are considered unreliable ( Barbour, 1912; Haas, 1950). In defense of Bleeker’s Sumatran record, one of us (JCM) has recently found a Sumatran snake that looks remarkably like Brachyorrhos but most likely does not belong in the genus. Similarly, the species has been reported from Borneo, and examination of one museum specimen labelled as Brachyorrhos albus (SMNS 2749) collected in Borneo in 1897 by H. Rolle, was in fact a bleached specimen of Calamaria cf schlegelii.
Brachyorrhos was reported twice from Timor in the 19 th century.A Timor specimen supposedly in the Leiden museum was probably collected by Müller and described by Günther (1858) and possibly also by Boulenger (1893). Müller (1857) reported Calamaria brachyorrhos (= B. albus ) on Ambon, but not Timor; this specimen was not found in the Leiden museum collection. We found this specimen in the British Museum (BMNH 442222) and doubt that it originated from Timor. However a second specimen (RMNH 576), also reported from Timor, was located in the Leiden museum. This specimen, collected by E. W. A. Ludeking in 1863, has several lines of evidence suggesting it originated in Seram. Data with Ludeking’s specimen reported “Papoetikbaai” [= Paputik Bay], Timor. However, three specimens of Draco lineatus bourouniensis were collected by Ludeking in 1863 on Seram at “Papoetik bay” ( Musters, 1983), as well as one specimen of Varanus cerambonensis ( Philipp et al., 1999) . The type location for the V. cerambonensis is Seram. Old maps of Seram leave only one option for the locality “Paputik Bay”, it is now known as Elpaputih Bay in Southwest Seram. Additionally, Kaiser et al. (2011) failed to find Brachyorrhos in the Timor-Leste herpetofauna. In our cluster analysis and PCA, BMNH 442222 associates with the Ambon population; and RMNH 576 associates with the Seram population.
There is a single specimen from the Aru Islands (MSNG 56342), received in 1873 by Beccari. In our cluster analysis and PCA this specimen associates closely with the Buru species, it is identical to the Buru species in almost every detail, although it has one primary temporal (most Buru specimens have two). Its presence in the Aru Islands seems improbable but cannot be completely ruled out.
Given the work of How and Kitchener (1997) and De Lang (2011) in the Lesser Sundas, Steubing and Inger (1999) in Borneo, and Hoesel (1959) in Java all of these localities are highly improbable for Brachyorrhos , and in the absence of voucher specimens its presence on these islands should not be assumed.
Reports of Brachyorrhos in Western Papua are based on members of the genus Calamophis Meyer. Therefore , Brachyorrhos is endemic to the Moluccas based on current information and our analysis suggests it is polytypic, containing at least four species. Note added in proof. Since this writing specimens have been collected at Kofiau Lake, Kopiau, Raja Ampat Islands, Papua (01°09'09.2''S, 129°51'42.5''E).
Results from a cluster analysis and PCA prompted a closer examination of the six specimens from Papua. They form a group morphologically distinct and geographically disjunct from the other 58 specimens examined. Fig. 3 View Fig illustrates the known distribution of Brachyorrhos and the related genus Calamophis .
Etymology. — Brachyorrhos is derived from the Greek words brachy = short, and orrhos = rump or tail; the gender is masculine.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Brachyorrhos Kuhl
| Murphy, John C., Mumpuni, Lang, Ruud de, Gower, David J. & Sanders, Kate L. 2012 |
Oxyorrhos
| Fischer, J 1879: 89 |
Rabdion
| Bleeker, P 1860: 42 |
Brachyorrhus
| Schlegel, H 1837: 25 |
Brachyorrhos
| Schlegel, H 1826: 236 |