
Protaphorura borinensis, Parimuchová, Andrea & Kováč, Ľubomír, 2016
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4098.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:4907385B-950F-44FB-8D0B-403947FD753E |
|
DOI |
https://doi.org/10.5281/zenodo.6089792 |
|
persistent identifier |
https://treatment.plazi.org/id/E55F9220-FFC9-A830-FF7C-A3DAFC1CFBD2 |
|
treatment provided by |
Plazi |
|
scientific name |
Protaphorura borinensis |
| status |
sp. nov. |
Protaphorura borinensis sp. nov.
( Fig.1–13 View FIGURE 1 – 5. P View FIGURE 6 – 9. P View FIGURE 10 – 13. P )
Diagnosis. PAO with 36–43 simple vesicles. Pso formula dorsally: 33/023/33343(4), ventrally: 1/000/00000. Subcoxae I with 1,1,1 pso each. Coarse granulation on head with up to 8 granules between p1–p1 chaetae. Abd. I– II with s and s' chaetae, s' always missing on Abd. III. Abd. V with chaeta s closest to pso c ( Fig. 8 View FIGURE 6 – 9. P ). Verticil C of each Tita with 3 chaetae. On Abd. I pso a and b placed between macrochaetae p2 and p5. VT with 11–15 distal and 2–3 basal chaetae on each side.
Type material. Holotype female and 14 paratypes (13 females, 1 male: Slovakia, Little Carpathians, Sedmička Cave, Veľký dóm Hall, collected by pitfall traps, 19.vi.–28.xi. 2014, leg. A. Mock, P. Ľuptáčik, A. Parimuchová.
Other material: Slovakia, Little Carpathians, Sedmička Cave, 6 females and 4 males, collected on walls and surface of water puddles on the cave sediment, 19.vi. 2014, leg. A. Parimuchová.
Description. Body length from 2.7 mm (males) to 3.3 mm (females), body shape typical for the genus, with anal spines on distinct papillae ( Fig. 1 View FIGURE 1 – 5. P ). Colour white alive and in ethyl alcohol. Granulation distinct and coarse especially on head, with up to 8 granules between p1 chaetae on hind margin of head ( Fig. 9 View FIGURE 6 – 9. P ).
Antennae equally long as head, area antennalis well marked. Ant. I with 11–14 chaetae (mostly 12). AOIII with 5 guard chaetae, 5 papillae, 2 smooth sensory rods shorter than papillae and 2 morel-like sensory clubs of equal size. Microsensillum in ventro-lateral position, at level of last guard chaeta ( Fig. 6 View FIGURE 6 – 9. P a, b). Subapical organite of Ant. IV placed in unprotected cavity. Ms on Ant. IV in ventro-lateral position, in 1/3 of segment length measured from base.
PAO with 36–43 simple vesicles ( Fig. 3 View FIGURE 1 – 5. P ). Labial palp of type A with mostly 7 (6–8) proximal chaetae ( Fig. 10 View FIGURE 10 – 13. P ). Basomedian field of labium with 4+4 chaetae, 4+4 postlabial chaetae in symmetrical arrangement. Maxillary outer lobe with simple palp, 1 basal and 2 sublobal chaetae ( Fig. 2 View FIGURE 1 – 5. P ). Pso formula dorsally: 33/023/33343(4), ventrally: 1/ 000/00000. Subcoxae I–III with 1 pso each. Ventral psx formula: 1/000/11110, femur and subcoxae I with 1 psx each, femoral psx sometimes open and visible as larger area of ungranulated cuticle ( Fig. 13 View FIGURE 10 – 13. P ). 1 pseudoporus on each subcoxa II, pseudopora on Th. I–III and Abd. II present but hardly visible.
Dorsal chaetotaxy plurichaetotic, on Th. I variable, chaetotaxy type from i 3 m to 2 i3 (2)m with modifications, mostly with 19–25 chaetae per half side of tergum ( Fig. 10 View FIGURE 10 – 13. P ). Chaetae s and s' on Abd. I and Abd. II present, s' seldom missing. On Abd. I–II pso a and b located between macrochaetae p2 and p5 ( Fig. 7 View FIGURE 6 – 9. P ). Abd. III without s', only pso a between macrochaetae. Abd. V always with chaeta s closest to pso c, and mostly also with chaeta s' and s" ( Fig. 8 View FIGURE 6 – 9. P ); ratio of chaetae M/ s= 19/8. Straight lines passing through the bases of K and K’ prespinal chaetae distinctly convergent; chaeta K x sometimes present ( Fig.12 View FIGURE 10 – 13. P ).
Furca remnant with distinct, arched cuticular fold with 2+2 chaetae in two rows (1+1 placed on fold, 1+1 posterior to fold), 4 chaetae in ma row, 4–5 chaetae in mm row and 5–7 chaetae in mp row ( Fig. 5 View FIGURE 1 – 5. P ). Male ventral organ absent. VT with 11–15 distal and 2–3 basal chaetae on each side.
Legs. Tita of all legs with 11 chaetae in distal verticil (A+T), 7 chaetae and chaeta M in verticil B and 3 (seldom 4) chaetae in verticil C. Proximal chaeta Y (placed above verticil C) present. Claw with 2 lateral teeth and 1 small inner tooth in the middle of claw length ( Fig. 4 View FIGURE 1 – 5. P ). Ratio length/ width of claw ~2.0.
Etymology. The species is named after the Borinský kras Karst, a small karst area near Bratislava, where the type locality (Sedmička Cave) is situated.
Biology. Specimens of P. borinensis sp. nov. were discovered in Sedmička Cave, Borinský kras Karst, in the Little Carpathians that is the westernmost mountain range of the Western Carpathians. The cave appears to be oligotrophic without active underground stream, bat guano or other deposited organic materials. Specimens were distributed in hind parts of the cave distant from the entrance, collected on walls and surface of the water pools. The new species shows obvious affinity to subterranean environment, up to date it is known only from type locality in spite of intensive sampling in nearby caves of the same karst area. P. borinensis sp. nov. displays troglomorphic traits, i.e. slightly elongated antennae and claws.
Distribution. Known only from type locality, Sedmička Cave, Little Carpathians, Slovakia.
Discussion. The new species is similar to the Holarctic P. ar m a t a in dorsal and ventral pso formula (33/023/ 33343, 1/000/00000) and distinctly convergent straight lines passing through the bases of prespinal chaetae. It differs from P. armata by larger body, higher number of vesicles in PAO and plurichaetotic Th. I tergum ( Table 1 View TABLE 1 ). The new species is the most similar to troglobiotic P. janosik Weiner, 1990 , species endemic to the Western Carpathians, in number of vesicles in PAO (~40) and strong plurichaetosis on Th. I tergum especially. However, macrochaeta p3, typical for P.janosik , is absent in the new species. Furthemore, P. borinensis sp. nov. differs from P. janosik by 11–15 distal chaetae on VT (18–22 in P. j a no s i k). Granulation, especially on head and Th. I, is more distinct in P. borinensis sp. nov. than in P. janosik ; only 8 granules are present between p1 chaetae in the new species compared to 12 in P. janosik (specimens from the type locality and several other populations from the Western Carpathians). Distinguishing between juveniles/subadults of P. janosik and P. borinensis sp. nov. can be problematic since subadult specimens of P. janosik often has indistinct macrochaeta p3 and lower number of granules between p1 chaetae. Anyway, cuticular fold of furca remnant in the new species is arched and well visible while it is very small and shallow in P. janosik Weiner, 1990 . Specimens of the new species and specimens of five different populations of P. janosik (incl. the population from the type locality) were sequenced by COI locus. Maximum likelihood tree placed P. janosik and P. borinensis population in two different branches with strong statistical support showing a clear separation of analysed specimens of P. janosik and P. borinensis sp. nov. (Parimuchová et al., unpubl.). Morphology and molecular variability of P. janosik populations from the Western Carpathian caves will be shown in the next paper. Thus, we assume P. borinensis , despite being morphologically similar to P. janosik , as a new species.
According to Pomorski (1990) the presence of chaeta s' on Abd. I affiliates the new species into common group together with P. s-vontoernei ( Gisin, 1957) and P. pseudovanderdrifti ( Gisin, 1957) . The same author, when evaluating variability of abdomin s' is missing. If s' is present, then both pso a and b are demarcated by macrochaetae. In the new species, there is a new combination: both pso a and b are set between macrochaetae and chaeta s' is absent. The new species is also similar to P. octopunctata ( Tullberg, 1876) in body length, length of antennae (equal to head), and variable tergal chaetotaxy with tendency to plurichaetosis. Nevertheless, there are specific characters separating the new species from others of “ octopunctata ” group characterized by area antennalis with 4 pso and parallel straight lines passing through the bases of prespinal chaetae. According to number of vesicles in PAO (~ 36), body length (~ 3 mm), 3 antero-dorsal and 3 postero-dorsal pso on head and presence of 2–3 pso on Th. II–III, we propose to assign the new species to “ multivesiculata ” group together with Protaphorura janosik , P. septempapillata and P. armata multituberculata .

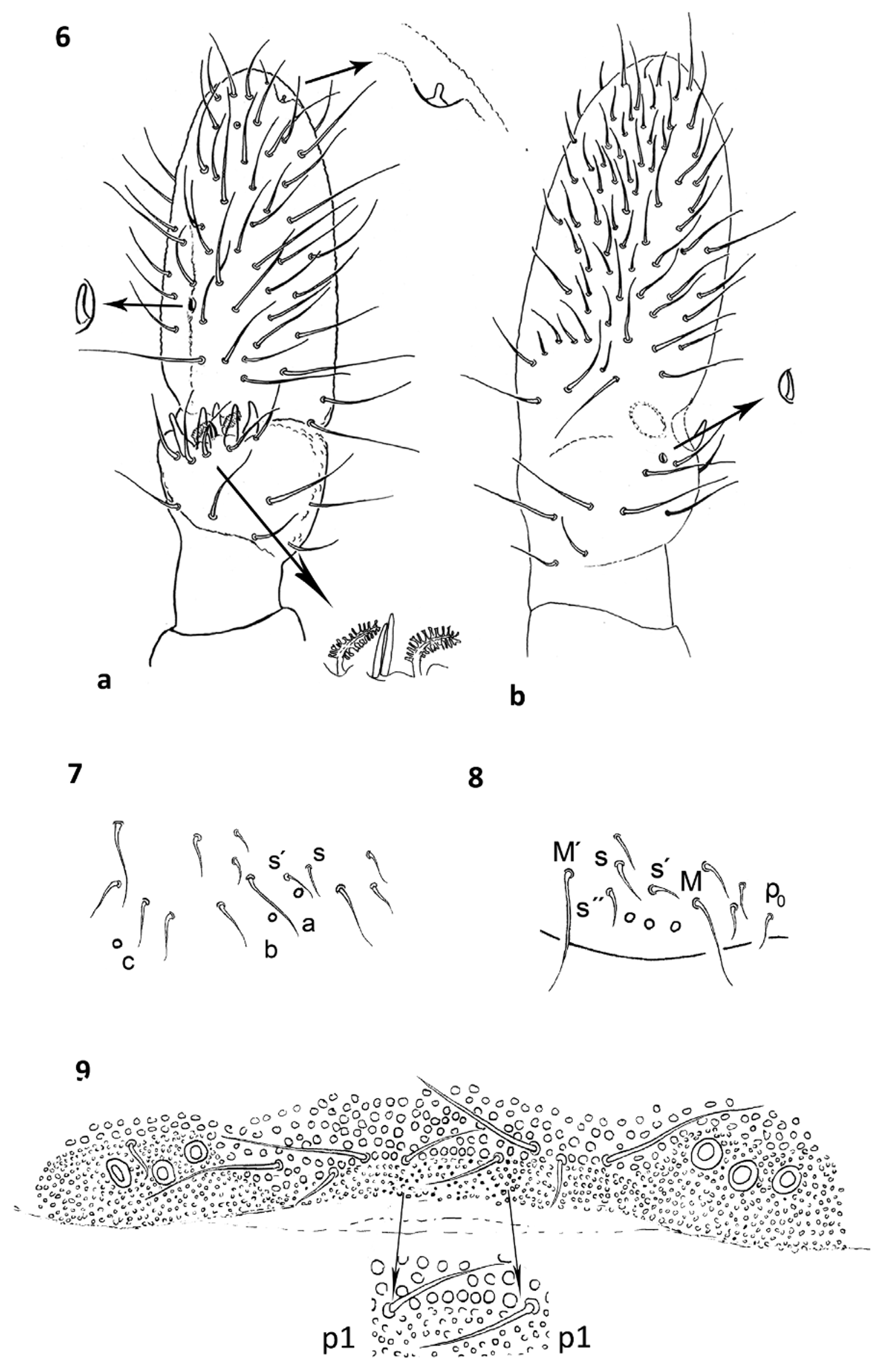

TABLE 1. Differential diagnosis of the species similar to P. borinensis sp. nov.
| Size (mm) PAO | Dorsal pso | Chaetotaxy of s´ on Th. I Abd. I | s´ on Abd. V | VT - distal chaetae | |
|---|---|---|---|---|---|
| P. janosik Weiner, 1990 | 2.9–4.3 36–58 | 33/022–3/33343 | plurichaetotic, +/- p3 as macrochaeta | + | 18–22 |
| P. tricampata Gisin, 1956 sensu Pomorski, 1998 | 1.8–2.0 28–35 | 33/023/33343 | i3-? | + | 8+8 |
| P. armata (Tullberg, 1869) sensu Gisin, 1952 | 1.5 28–35 | 33(4)/023/3334(5)3 | i3- - | - | 8+8 |
| P. a. multituberculata (Stach, 1934) | 4.2 54–64 | 32/022/33343 | i2m? | - | ? |
| P. borinensis sp. nov. | 2.7–3.3 36–43 | 33/023/33343(4) | plurichaetotic, + p3 as mesochaeta | + | 11–15 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.