
Mictini
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3641.5.60 |
|
publication LSID |
lsid:zoobank.org:pub:2F8F16B1-5D5C-4A31-94CD-0BD92EF728BE |
|
DOI |
https://doi.org/10.5281/zenodo.5658445 |
|
persistent identifier |
https://treatment.plazi.org/id/E57687B7-FF90-5C38-59C2-FAD5FF7AE58E |
|
treatment provided by |
Plazi |
|
scientific name |
Mictini |
| status |
|
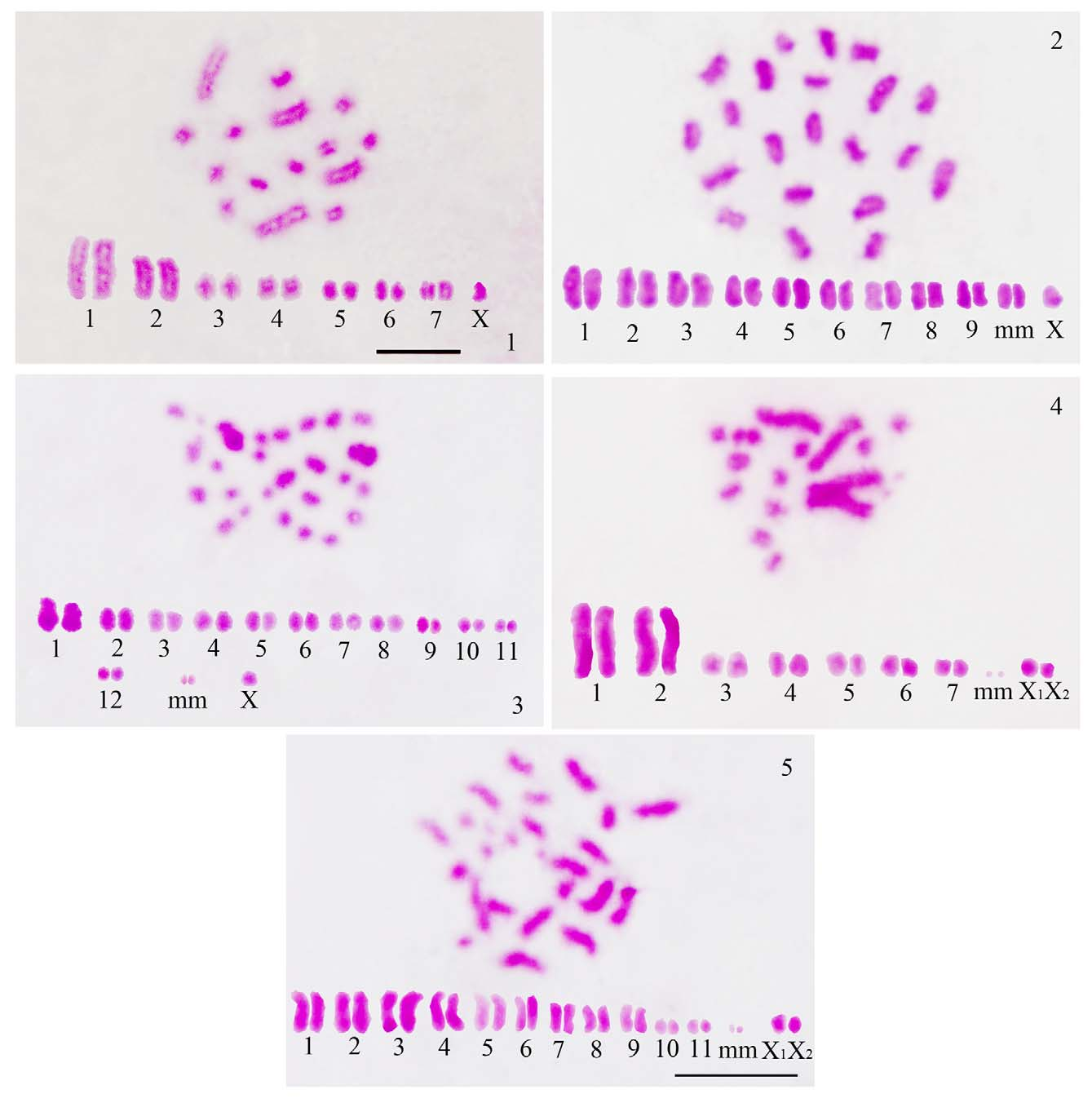
The tribe Mictini is found only in the Eastern Hemisphere and comprises 47 genera (Schuh & Slater 1995). Cytologically, this tribe is represented by 13 species belonging to 6 genera. The present study adds 3 species viz., Anoplocnemis binotata , Ochrochira nigrorufa and Prionolomia sp. to the pre-existing cytogenetic data for Mictini .
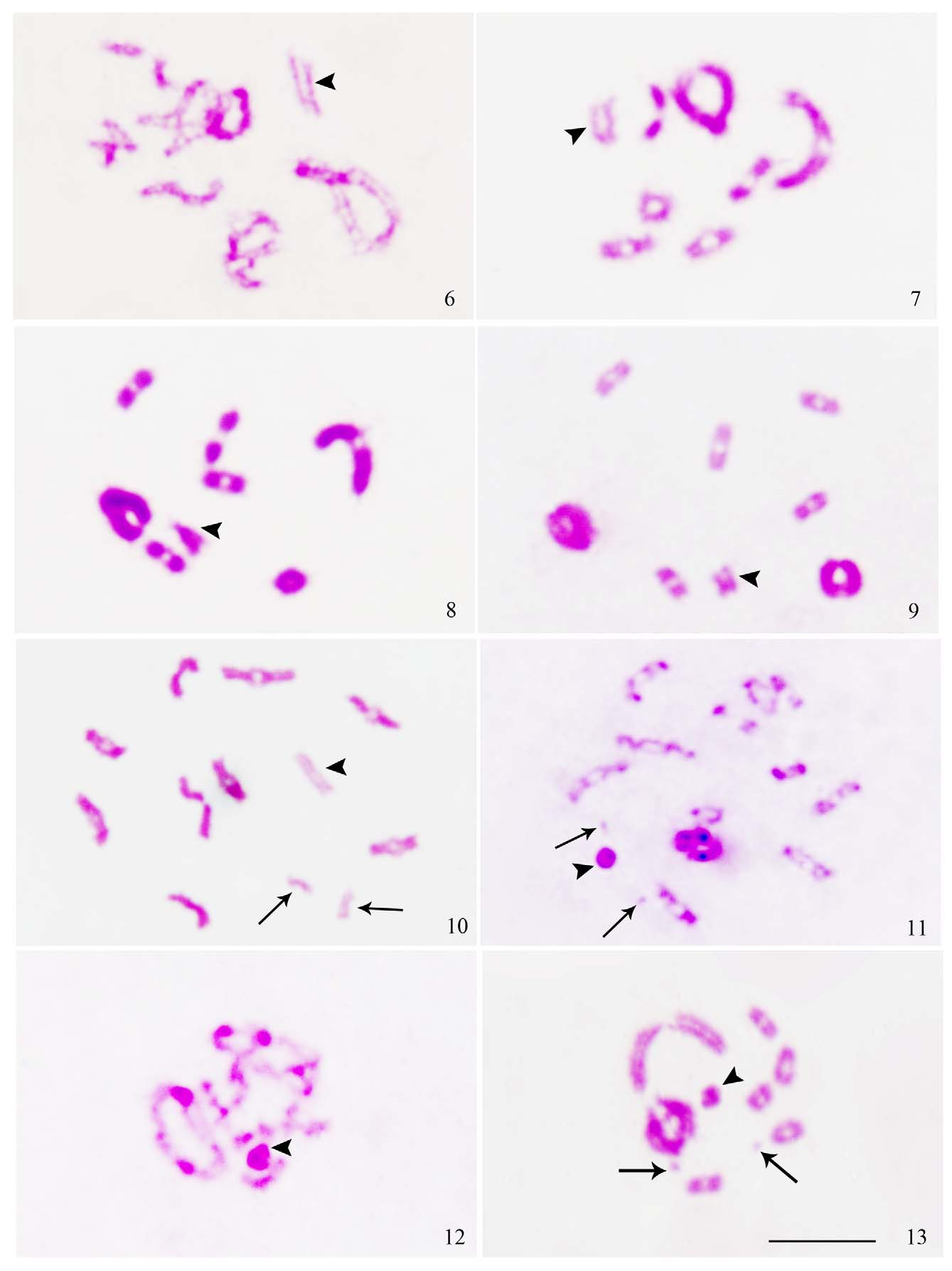
The diploid chromosome complement is 2n=15=14A+X0 in Anoplocnemis binotata as also recorded in Anoplocnemis phasiana (Sands 1982) and Anoplocnemis compressa (Kaur & Bansal 2012a). Two pairs of autosomes are extremely large and the rest of the 5 pairs are medium. Microchromosomes are absent ( Fig. 1 View FIGURES 1 – 5 ). X shows a characteristic behavior during diplotene. At the start of diplotene, X appears decondensed with chromatids lying parallel. But as diplotene progresses, it shows differential condensation along its length, being partly condensed and partly decondensed and appears tong-shaped, sometimes bent also. By late diplotene/diakinesis, condensation of the X is completed ( Figs. 6–9 View FIGURES 6 – 13 ). This observation led us to re-examine diplotene stages of Anoplocnemis compressa ( Figs. 14–17 View FIGURES 14 – 21 ) and Anoplocnemis phasiana ( Figs. 18–21 View FIGURES 14 – 21 ); a similar behavior was recorded in them also which, however, was overlooked by Sands (1982) in Anoplocnemis phasiana and was misinterpreted as X1X2 by us in Anoplocnemis compressa in an earlier study (Kaur & Bansal 2012a).
The diploid chromosome number of Ochrochira nigrorufa is 2n=21=18A+ 2m +X0, which is the same as in Ochrochira granulipes , Ochrochira rubrotincta and Ochrochira aberrans (Dey & Wangdi 1990; Sands 1982; Kaur & Bansal 2012a). However, in Ochrochira albiditarsis and Ochrochira sp., 2n is 21=20A+X0 and 26=22A+ 2m + X1X2 0, respectively as reported by Parshad (1957b) and Jande (1959). A gradual gradation in size has been observed in the autosomes. Microchromosomes in Ochrochira nigrorufa are unusually large as compared to other coreids ( Fig. 2 View FIGURES 1 – 5 ). Such large microchromosomes have earlier been recorded in Ochrochira granulipes , Ochrochira rubrotincta , Ochrochira aberrans and Elasmomia granulipes (Sands 1982; Dey & Wangdi 1990; Kaur & Bansal 2012a). These microchromosomes remain as univalents during diplotene, but get terminally associated to form a pseudobivalent in late prophase I and orientate in the center of the autosomal ring at metaphase I.
Prionolomia sp., cytologically studied for the first time, possesses a diploid chromosome complement of 2n=27=24A+ 2m +X0. One pair of autosomes is distinctly large, 7 pairs are medium and 4 pairs are distinctly small ( Fig. 3 View FIGURES 1 – 5 ).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Coreinae |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Coreinae |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Coreinae |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Coreinae |
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Coreinae |