
Pseudohydrosme gabunensis Engl.
|
publication ID |
https://doi.org/10.7717/peerj.10689 |
|
DOI |
https://doi.org/10.5281/zenodo.5847044 |
|
persistent identifier |
https://treatment.plazi.org/id/EF0FA647-3575-C240-FFB0-FA2BD63C5F7A |
|
treatment provided by |
Felipe |
|
scientific name |
Pseudohydrosme gabunensis Engl. |
| status |
|
2. Pseudohydrosme gabunensis Engl. View in CoL ( Engler, 1892: 455; Engler & Prantl, 1897: 59; Brown in Thistleton-Dyer, 1901: 161; Engler, 1911: 48; Bogner, 1981: 33; Hetterscheid & Bogner, 2013: 104–113)— Figs. 3 View Figure 3 and 5 View Figure 5 .
Holotype: Gabon, Estuaire Province, Libreville, Sibang , “Gabun, Mundagebiet ; Sibang-Farm am Ufer des Maveli ” fl. 13 October 1881, Soyaux 299 ( Holotype: B100165306 , Herbarium specimen, image!)
Terrestrial herb, rhizome light brown, ellipsoid or subcylindric, erect or oblique, 9–12 cm diam. to 15 cm long, surface with transverse ridges. Roots fleshy 5–8 mm thick, brownish yellow, sometimes developing new plants at their tips ( Hetterscheid & Bogner, 2013).
Leaf 1–1.3(–2.2) m tall, petiole terete, 1–1.4 cm diam. at base, dark green olive and spotted, with small yellowish white points; prickles 1–2 mm long. Blade of youngest seedlings sagittate-elliptic c. 5 cm long, 3–4 cm wide, basal sinus c. 2 cm long, breadth variable (see Hetterscheid & Bogner, 2013). Successively formed blades developing slits and divisions. Blade of mature leaves dracontoid, primary divisions 30–35 cm long, pinnatisect, lobes in each division 5–8, dimorphic, larger, distal lobes elliptic (4–)8–23 cm long, (2–)3–7(–11) cm, apex truncate, bifid, (0.5–)1–3 cm long; smaller, proximal leaflets ovate, 4.5–8 cm long, 2.5–5 cm wide, apex cuspidate; lateral veins 4–8 on each side of the midrib, conspicuous on abaxial surface, running to the margin or forming an incomplete submarginal nerve, higher order veins reticulate.
Inflorescence: Cataphylls 4–6, membranous, reddish white or brown-purple, slightly spotted, phyllotaxy spiral, proximal ones subtriangular shorter, distal ones becoming longer and oblong elliptic, towards the spathe 1.5–29 cm long, (1–)2–2.5 cm wide; peduncle (3–)5–9 cm long, 1–1.5 cm diam., colour as petiole, with minute sparse greenish white prickles 1–2 mm long. Spathe (30–)40–55(–70) cm long, fornicate, basal half (20–25 cm long) funnel-shaped to subcylindrical, fleshy and to 5 mm thick, limb comprising the distal half of the spathe, flaring widely and curving forward, the apex obtuse, margin undulate; outer surface uniform bright pale yellow, greenish yellow or yellow white; inner surface of blade mostly pale yellow or yellowish white, in an irregular marginal band, with a dark purple central area separated by an irregular margin down to the base of the tube; mouth facing horizontally, usually orbicular or elliptic. Spadix with “unpleasant smell, but not so strong as some Araceae ” ( Van der Laan 7641, WAG) or “faintly of lettuce” ( Hetterscheid & Bogner (2013)) or “of slightly bad cabbage” (David Prehsler, University of Vienna communication to Cheek November 2020), sessile, subcylindrical, (6–)9–12.5 cm long, (1.5–)2–2.5 cm diam. Female zone (2–)3.5(–4) cm long, female flowers completely covering the surface of the axis, usually contiguous with but constricted at the junction with the male zone. Male zone (3.5–)6–8.5 cm long, at base abruptly wider than the female zone, tapering to the rounded apex, completely covered in fertile male flowers. Sterile appendix absent.
Male flowers with 2–5 stamens, stamens densely packed, sometimes paired or in groups of 3 or 5, sessile, 4 mm long, in plan view isodiametric, subprismatic, 5–6- faceted, in cross section c. 1.8(–2) mm × 1.2 mm wide, apex convex purple, sides white, anther thecae c. 3 mm long, opening by an apical pore, pollen orange or yellow, in strings. Female flowers white with ovary yellowish-white globose or ellipsoid, 2–3 mm diam., 2(–3)-locular; style 1–1.5 mm long, 1.5 mm diam., stigma black to reddish brown, surface papillose, 2 mm wide, bilobed, lobes with a broad concave area, apex rounded.
Berry, thinly fleshy, transversely ellipsoid, laterally compressed, rarely globose, 0.8–1.2 cm long, 1.5–1.6 cm wide, white, ripening purple-black, surface wrinkled when ripe, style and stigma persistent, (1–)2-seeded, apex rounded-truncate, base stipitate, stipes (2–) 3–4 mm long, c. 2 mm diam. Seeds subglobose to broadly ellipsoid, one side flattened, the other convex, 9 mm long, 7 mm wide.
Phenology: flowering in the wild mid-September–late October.
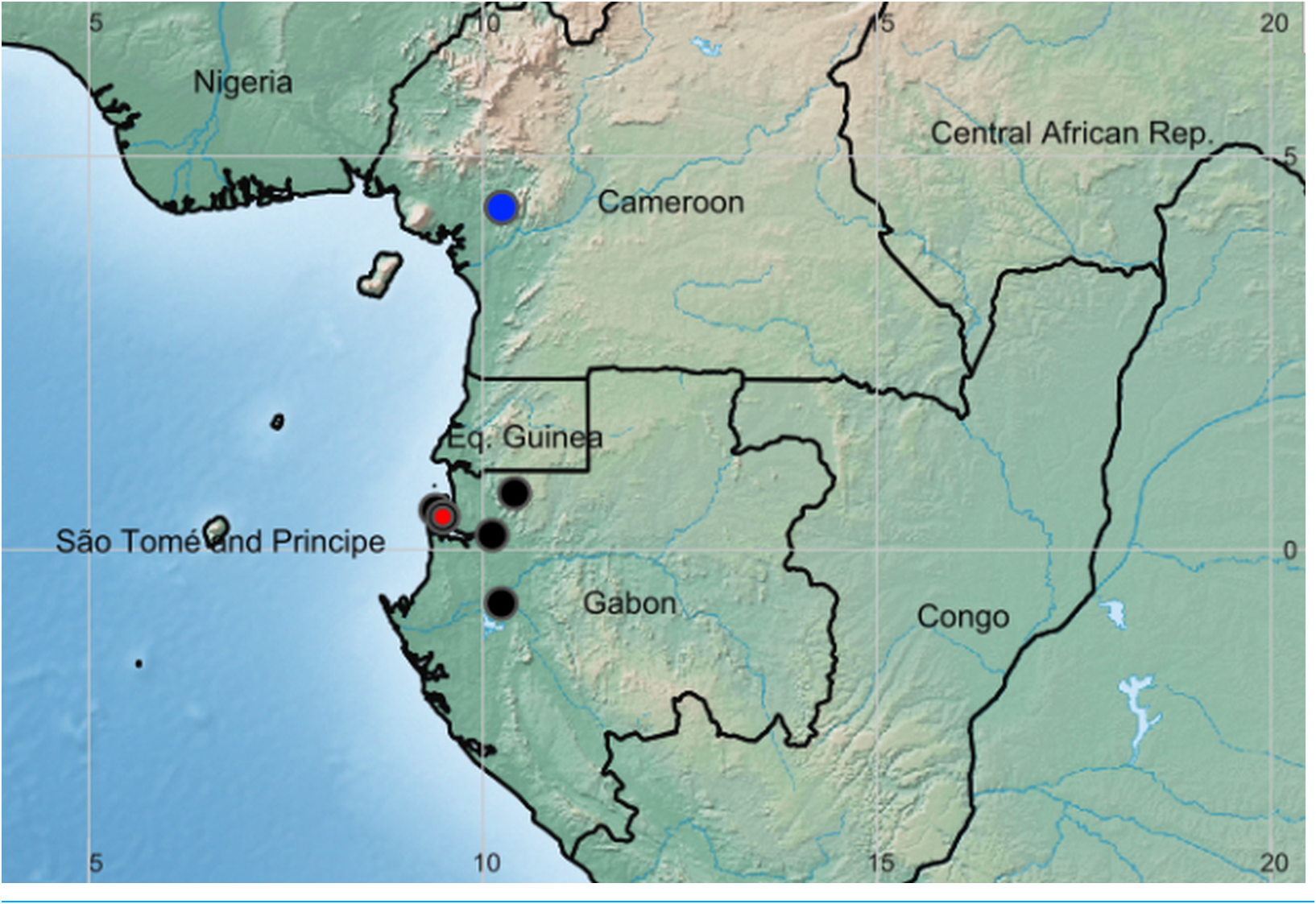
Distribution and ecology: Gabon, Estuaire, Moyen-Ogooué (probably) and Woleu-Ntem Provinces, known from five sites in lowland rainforest sometimes with Aucoumea gabonensis (Burseraceae); 0–100 m alt. Possibly also in Congo (location unknown, see notes below).
Etymology: meaning “coming from Gabon” (formerly, in German “Gabun”).
Local names and uses: none known.
Additional specimens: Gabon, Woleu-Ntem Province, c. 15 km NE Asok, 600–700 m alt., (leg. Breteler and De Wilde s.n. 21 August 1978) cult. Wageningen, fl. 13 March 1984, Van der Laan 764 (Bot. Gard. No. 978PTGA550), WAG0351246, WAG0351247 images!) ; Estuaire Province, Libreville, Sibang: “Sibang”, hinter der Station forêstier ; wächst im sandigen Lehmboden im Regenwald, sehr schattig, c. 20 m, fl. 29 October 1973, Bogner 664 (K!, M n.v. US n.v.) ; Sibang, st. 10 April 1994, Wieringa and Haegens 2710 (WAG0181636, WAG0181637 images!) ; Sibang Forest, st. 1 Dec. 1994, Wieringa 3308 (WAG0181631, WAG0181632, WAG 0181633, WAG0181634, WAG0181635 images!) ; Sibang Arboretum fl. 25 Oct. 2005, Sosef et al. 2029 (WAG 0223594, WAG0223595 images!, WAG8004057, WAG0108030, WAG.1665445) ; Kango, plantations de Assouko, près de poste de Kango, le Komo (estimated as 0 Ǫ 10′ 41.8″ N, 10 Ǫ 06′ 45.54″ E) , fl. 2 Oct. 1912, Chevalier 26828 (P02093245 image!) ; Forêt de la Mondah, road from Libreville to Santa Clara, fl. 16 Sept. 1981, Breteler, Lemmens, Nzabi 7772 (WAG0449339, WAG0449339, WAG0449340 images!) ; St. Clara, Tussen ± 50–100 m, Linkerkant, Zij-pod naar St. Clara, sterile, no date, Breteler s.n. (WAG044938, image!) ; Moyen-Ogooué Province: “Congo français”. Ogooué (estimated as 0 Ǫ 41′ 18″ S, 10 Ǫ 13′ 55″ E), fl. 1894–95, Leroy 23 (PO2093240, PO2093241 images! two sheets) .
Cultivated in Europe exact source unknown: ex Gabon, probably Sibang, fl. April 2012, leg. Bogner 3006 (BR0000019808871, image!).
Those specimens listed above which are sterile, for example Wieringa 3308 (voucher for DNA studies of the genus, see above), Wieringa and Haegens 2710, are only provisionally identified as P. gabunensis . It is possible that these specimens might belong to another species of the genus (although unlikely since these specimens were collected at Sibang Arboretum where in recent years only this species of the genus has been collected in flower). Equally they may even represent a species of the genus Anchomanes .
Conservation: Pseudohydrosme gabunensis is possibly extinct at some of its historical locations and is threatened at all of those which remain. At the type location, Sibang, formerly far outside Libreville, at least four gatherings have been made in what is now a small and highly visited forest patch inside Libreville (see notes under P. buettneri above). Measured on Google Earth, the forest is approximately a square, c. 470 m N to S and 420 m W to E, or about 0.25 km 2 (grid reference: 0 Ǫ 25′ 56.05″ N, 9 Ǫ 29′ 23.64″ E, 49 m alt.). It is now completely surrounded by the dense urban settlement of Libreville which has expanded greatly in the last 60 years. In 1960, at independence, the population of Libreville was 32,000. Since then it has expanded 20-fold to, in 2013, 703,904
(https://en.wikipedia.org/wiki/Libreville, accessed 19 September 2020) and has a vastly greater footprint. Sibang Arboretum, the surviving patch of forest of a once much greater area, is now known as one of the top two tourist destinations in Libreville.
At the Cap Santa Clara location, the Forêt de la Mondah, known since 2012 as the Raponda Walker Arboretum ( Walters et al., 2016), two collections were made, one in 1981 (see additional specimens). Since created as a protected area in 1934, it has been reduced in size, losing 40% of its area in 80 years to habitat clearance and degradation due to its close proximity (c. 15 km) to the metropolis of Libreville which draws upon its trees for timber and firewood ( Walters et al., 2016). It is not clear if either of the two specimens from St. Clara were from within the current protected area.
The species has not been recorded from the Ogouué River since it was collected there by Leroy (1894–1895), despite intensive recent surveys in the lower reaches of the river whence it was probably collected. We have georeferenced the Leroy record on Lambarene since in Leroy’ s time this was a trading post on the lower reaches of the river and it is credible that he stopped and collected there, but this is uncertain. The historic site on the Komo River at Kango, whence it was collected by Chevalier (26828, P; fl. 2 Oct. 1912) is now on a major transnational route, and on Google Earth shows multiple cleared areas due to development. It is possible that it no longer survives at this location, especially since it has not been recorded here or anywhere near, in a century, despite the peak decades of botanical collection in Gabon having been at the end of the 20th century ( Sosef et al., 2005). Pseudohydrosme gabunensis was assessed as Endangered, EN B2ab (ii, iii) by Lovell & Cheek (2020) since it is or was known from ten specimens at five locations globally, with an area of occupation estimated as 24 km 2 using the 4 km 2 cell sizes preferred by IUCN (2012) and the threats detailed above. Threats in the Libreville area have already resulted in the possible global extinction of nine species, including Pseudohydrosme buettneri (see under that species, above). The extent of occurrence is calculated as 4,150 km 2. If the identification of the Congolese specimen can be completely confirmed as this species, and the site of its collection discovered, the area of occupation will likely be increased to 28 km 2 and the extent of occurrence also increased.
Notes. The location given in the protologue for the type specimen (see above) is similar to that of Pseudohydrosme buettneri but more detailed. The Munda is the estuary that forms the eastern edge of the peninsula on which Libreville sits. Tributaries of the Munda drain the Sibang area, one of which may have been known as the Maveli, on the forested banks of which Soyaux recorded collecting the type of Pseudohydrosme gabunensis .
Herman Soyaux is reported to have collected herbarium specimens from Loango in Gabon from 1875 to 1882 ( Anon, 1901).
The specimens Leroy 23 and Chevalier 26828 (both P) had been determined as Amorphophallus until identified by Bogner (M) as Pseudohydrosme gabunensis in Dec.
2012. In contrast, Wieringa 4358 (WAG) determined as this species, and cited as such in Sosef et al. (2005) is in fact an Amorphophallus , evident in the larger leaf blade divisions all being acuminate not bifid, and the tuber being described as having the roots from the top (vs.scattered along the length). Similarly, Wieringa 3308 (WAG), correctly cited in Sosef et al. (2005) as Pseudohydrosme gabunensis , was originally collected as an Anchomanes until determined by Hetterscheid in April 1996. Van der Laan 764 (WAG) had been identified as Anchomanes nigritianus Rendle until redetermined by Bogner in September 2012.
Pseudohydrosme gabunensis is the most common and widespread member of the genus. However, it is still extremely rare and with a highly restricted range in the wild.
It is sought after by private collectors of aroids and live rootstocks and seed attract high prices on the internet. Fortunately, it is found in several large public botanic gardens including in Australia, Germany, France, Netherlands, UK and USA. We believe that plants are probably not collected from the wild (but this cannot be ruled out), rather they are propagated from those already in cultivation, probably from seed derived from the Netherlands.
The collection reported in Hetterscheid & Bogner (2013) as from Congo must be treated with caution. Since it is greatly disjunct (at least c. 300 km) from the known range of this species, it may even represent a further new species. We have not been able to view this specimen. However, Hay (in litt.) states that he has seen photographs of flowering material and that it looks extremely like P. gabunensis in terms of spathe shape and colouration, therefore this is the most likely identification. The specimen concerned should be located, the spadix studied carefully to determine the species beyond doubt, and an attempt made to rediscover the source population.
Differences between Pseudohydrosme gabunensis and P. buettneri are detailed under the last species. There is no doubt that Pseudohydrosme gabunensis is much more closely similar to P. ebo than to P. buettneri . However, the larger size of the spathes in P. gabunensis , their different colour and patterning, the usually bilobed style and bilocular female flowers densely covering the axis, all serve, together with the vegetative characters, to separate it from P. ebo (see also Table 1 View Table 1 below).
Floral visitors: Bogner (1981) collected as inferred pollinators two different flies identified as Diptera: Choridae, Sphaeroceridae, and two different beetles identified as Coleoptera: Scaphidiidae, Staphylinidae in association with Bogner 664.
Reproductive biology: Hetterscheid & Bogner (2013: 106) working with cultivated plants, report that the female flowering phase is indicated by a faint yet clear lettuce-like scent as the spathe opens, at which time, for 2 days, the receptive stigmas are wet and sticky. After this time the stigmas turn darker brown, desiccate and are no longer receptive. Individuals are obligate outcrossers. Fruits take up 10 months to mature ( Hetterscheid & Bogner, 2013).
Germination and development: Germination takes 3 weeks to 10 months, producing a single small sagittate, entire leaf from a small rhizome. For several months to two years, new leaves are produced consecutively, usually each larger than its predecessor ( Hetterscheid & Bogner, 2013). From the second leaf onwards slits may develop in the blade, and within two years the successively produced blades first becomes divided and finally develop the mature dracontoid pattern (see description). First flowering has occurred in as little as five years from first sowing ( Hetterscheid & Bogner, 2013). In the wild, the time to maturity is likely to take longer due to predation, competition, and likely lower availability of nutrients

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
|
Section |
Pseudohydrosme |