
Leptalpheus cf. forceps Williams, 1965
|
publication ID |
https://doi.org/10.5281/zenodo.4689970 |
|
DOI |
https://doi.org/10.5281/zenodo.4893502 |
|
persistent identifier |
https://treatment.plazi.org/id/EF3D87DD-FFFB-5436-FF1C-FB5529555E49 |
|
treatment provided by |
Felipe |
|
scientific name |
Leptalpheus cf. forceps Williams, 1965 |
| status |
|
Leptalpheus cf. forceps Williams, 1965 View in CoL
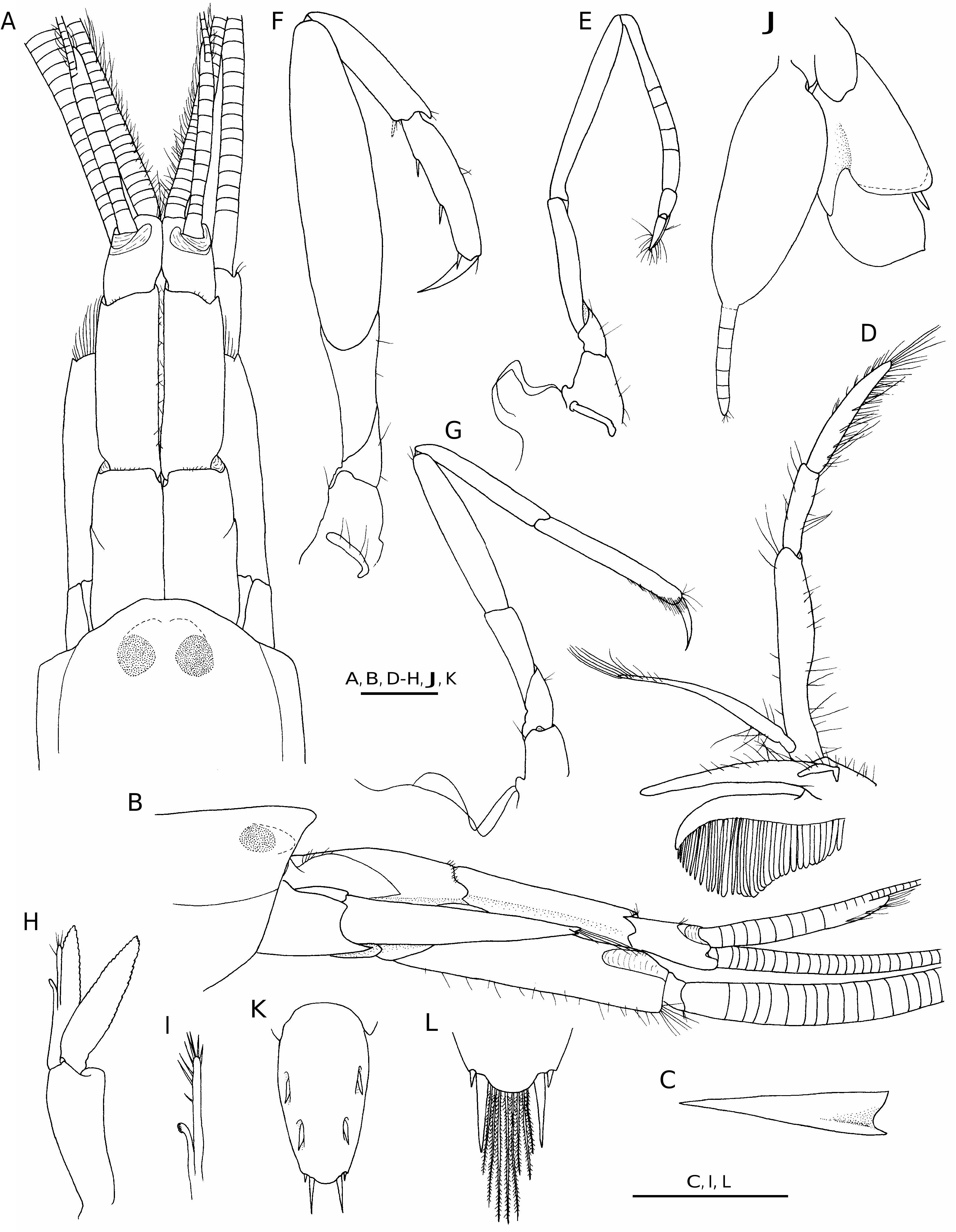
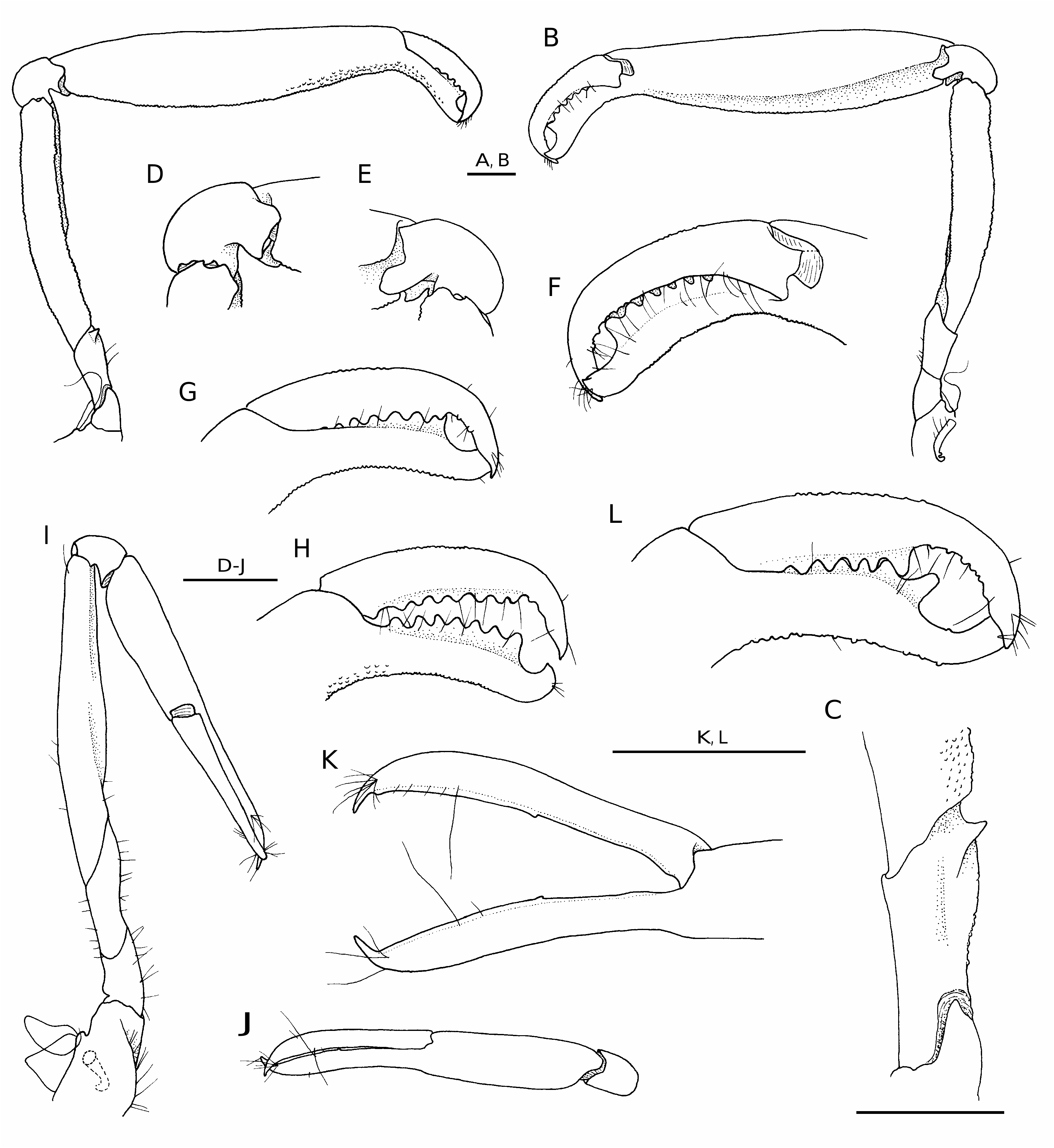
( Figs 4 View FIG ; 5 View FIG ; 6A, B View FIG )
Leptalpheus forceps Williams, 1965: 194 View in CoL ; 1984: 101. — Dawson 1967: 224. — Saloman 1971: 67. — Chace 1972: 77. — Abele & Kim 1986: 194, 228. — Felder & Rodrigues 1993: 366. — Holthuis 1993: 203. — Anker et al. 2006b: 686 View Cited Treatment .
(?) Leptalpheus forceps View in CoL – Christoffersen 1980: 135; 1998: 361. — Hermoso-Salazar 2001: 1278.
MATERIAL EXAMINED. — Costa Rica. Caribbean coast, Cahuita, Punta Uva, small mangrove river, coarse sand, from burrow of Lepidophthalmus richardi (host fcn 05- 118, will be deposited in MNHN), bait suction pump, depth 10-30 cm, coll. A. Anker, I. Wehrtmann and L. Harris, 26.XI.2005, fcn 05-119, 1 ♂, CL 8.2, TL 22.2 (MNHN-Na 17066). — Same collection data, from burrow of Lepidophthalmus richardi (host fcn 05-120, UCRMZ 2420-02 ), fcn 05-121, 1 ♂, CL 6.1, TL 15.3 ( UCRMZ 2420-01 ) ; 1 specimen (CL and TL not measured, sex not determined) preserved in Ambion RNA-later for molecular studies (fcn 05-006).
DESCRIPTION For complete description see Williams (1965, 1984). Detailed drawings of Cahuita specimens are provided in Figures 4 View FIG and 5 View FIG .
Size
The two Costa Rican male specimens were 6.1 mm CL, 15.3 mm TL, and 8.2 mm, 22.2 mm TL. Williams’ (1965) type specimens were 7.8 mm CL (female holotype) and 5.2 mm CL (male paratype).
Colour pattern
Semitransparent with yellowish and reddish chromatophores, more intense on tail fan, antennular peduncles,eyestalk and along posterior margins of the carapace and abdominal somites, forming transverse bands; some pale blue and greenish chromatophores present on the antennal scaphocerite and antennular peduncle; merus and carpus of major cheliped with reddish and some bluish chromatophores, major chela hyaline-whitish. Th e above-described pattern is much less conspicuous on a dark background: the shrimp appears as uniform whitish, with pale reddish bands on the abdomen and tail fan (see black-and-white photographs in Figure 6A, B View FIG ). The specimens from Fort Pierce, Florida (previously deposited in the MNHN) had pale greenish transverse bands on the abdomen; the antennules and the antennae were pale lettuce-greenish; the tail fan had locally pinkish, bluish and greenish tinge ( Anker et al. 2006b). Williams (1984) described the colour of L. forceps as “translucent, colorless in life; eggs light green in formalin”.
ECOLOGY
The present specimens were collected from burrows along the sea-side of a small river estuary, about 5 m wide, fringed with mangrove.Two specimens were collected together with their hosts, the callianassid ghostshrimp Lepidophthalmus richardi Felder &
A
Manning, 1997 ( Fig. 6C View FIG ). Elsewhere, L. forceps was found in burrows of the present host’s congeners, L. siriboia Felder & Rodrigues, 1993 ( Christoffersen 1980, as Callianassa jamaicensis ) and L. louisianensis (Schmitt, 1935) ( Felder & Rodrigues 1993) , as well as the upogebiid mudshrimp, Upogebia affinis
(Say, 1818) ( Williams 1965). Williams (1965, 1984) provided a brief summary of the biology of L. forceps .
HOST
Lepidopthalmus richardi ( Fig. 6C View FIG ) was previously known only from the type locality in Belize ( Felder & Manning 1997). Th e record from Cahuita represents a second finding of this species and a considerable extension of its distribution range from Belize southward to Costa Rica.
REMARKS
The two male specimens from Costa Rica ( Figs 4 View FIG ; 5 View FIG ) agree reasonably well with the illustrated female holotype from North Carolina (cf. Williams 1965), except for the more robust antennular peduncles (compare Fig. 4 A View FIG and Williams 1965: fig. 1B); the slightly shorter scaphocerite, reaching to about 2/3 length of the second segment of the antennular peduncle ( Fig. 4A View FIG ) vs. reaching almost to the end of this segment in the type ( Williams 1965: fig. 1B); and the more elongate lateral plate on the coxa of the third maxilliped (compare Fig. 4D View FIG and Williams 1965: fig. 2F). Williams (1965) noted that “the fingers of the major chela are slightly less gaping in females than in males” and that “there is a slight variation in the number of teeth on the fingers of the major cheliped”, however, without providing the exact range of this variation. Th e dentition of the type includes six teeth ( Williams 1965: fig.1G), whereas the two examined Costa Rican specimens have six and seven teeth, respectively (not counting the small most proximal tooth visible only when chela is widely opened) ( Fig. 5 View FIG F-H, L). Th e most distal tooth on the pollex may be more protruding than the others ( Fig. 5L View FIG ). However, the most important difference between the specimens from Costa Rica and North Carolina lies in the development of a segmented appendix on each uropodal endopod. These appendices (caudal filaments) are present in both males from Cahuita ( Fig. 4J View FIG ), and absent in the female holotype and apparently also in all paratypes of L. forceps ( Williams 1965: fig. 2L). Similar caudal filaments were observed in one male specimen of L. felderi from Isla Margarita, Venezuela ( Anker et al. 2006b: fig. 5C); however, in other males and in females of this species, the distal margin of the uropodal endopod was rounded. Th e function of these structures, if there is any, as well as their occurrence in some individuals and absence in others remain unexplained. Thus L. felderi appears to be a highly variable species, but more specimens and DNA sequencing are needed to determine the full extension and the genetic base of this variation, and to decide whether L. cf. felderi from Costa Rica is conspecific with L. felderi from North Carolina and Florida.

| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Leptalpheus cf. forceps Williams, 1965
| Anker, Arthur 2008 |
Leptalpheus forceps
| ANKER A. & VERA CARIPE J. A. & LIRA C. 2006: 686 |
| FELDER D. L. & RODRIGUES S. A. 1993: 366 |
| HOLTHUIS L. B. 1993: 203 |
| ABELE L. G. & KIM W. 1986: 194 |
| WILLIAMS A. B. 1984: 101 |
| CHACE F. A. JR. 1972: 77 |
| SALOMAN C. H. 1971: 67 |
| DAWSON C. E. 1967: 224 |
| WILLIAMS A. B. 1965: 194 |