
Isophya hospodar (Saussure, 1898)
|
publication ID |
https://doi.org/10.11646/zootaxa.3658.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:C02D1C74-25C0-41DD-B098-62098EB7B62A |
|
DOI |
https://doi.org/10.5281/zenodo.5617313 |
|
persistent identifier |
https://treatment.plazi.org/id/F26F3128-3900-FF98-B1B0-0CDEFE4E9B99 |
|
treatment provided by |
Plazi |
|
scientific name |
Isophya hospodar (Saussure, 1898) |
| status |
|
Isophya hospodar (Saussure, 1898)
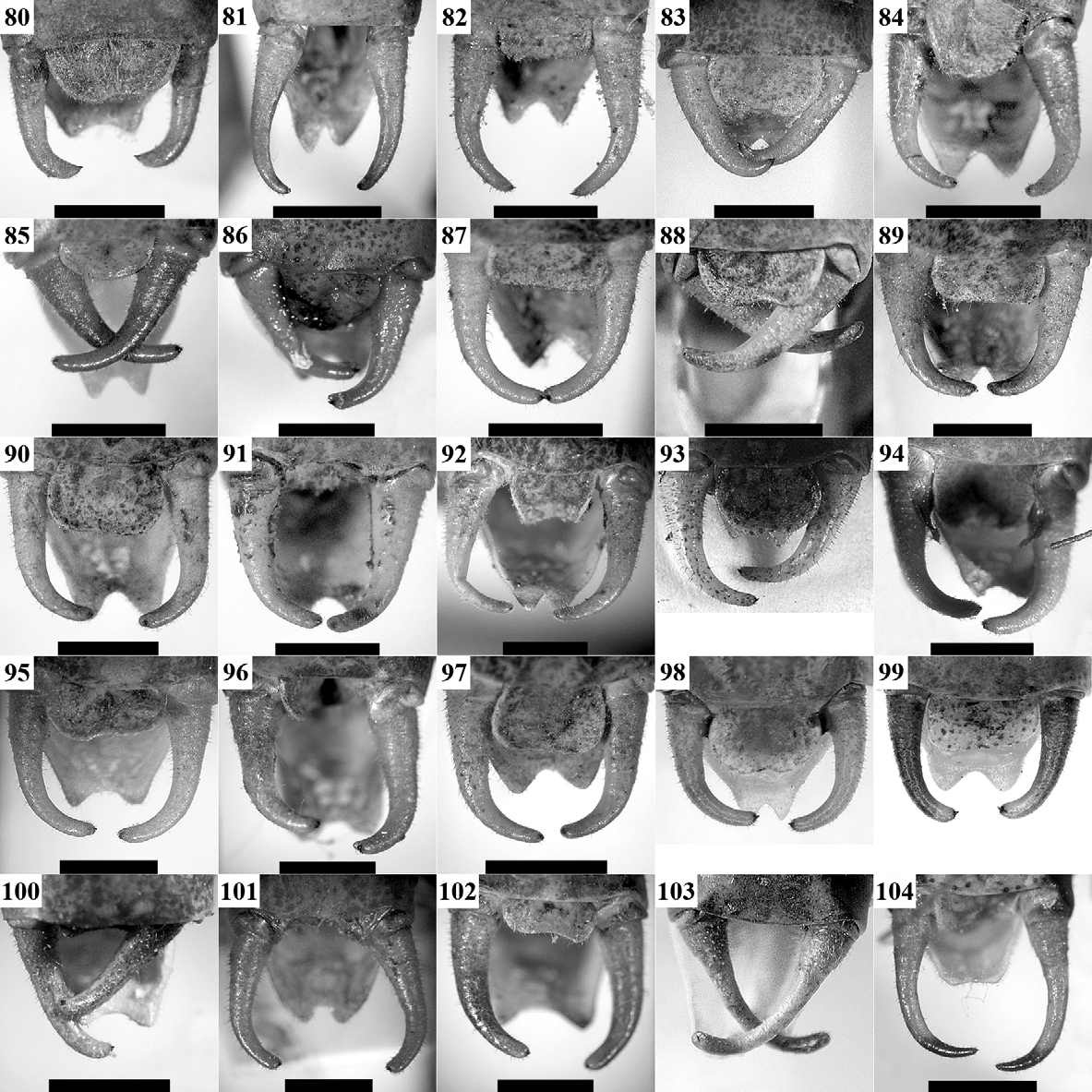
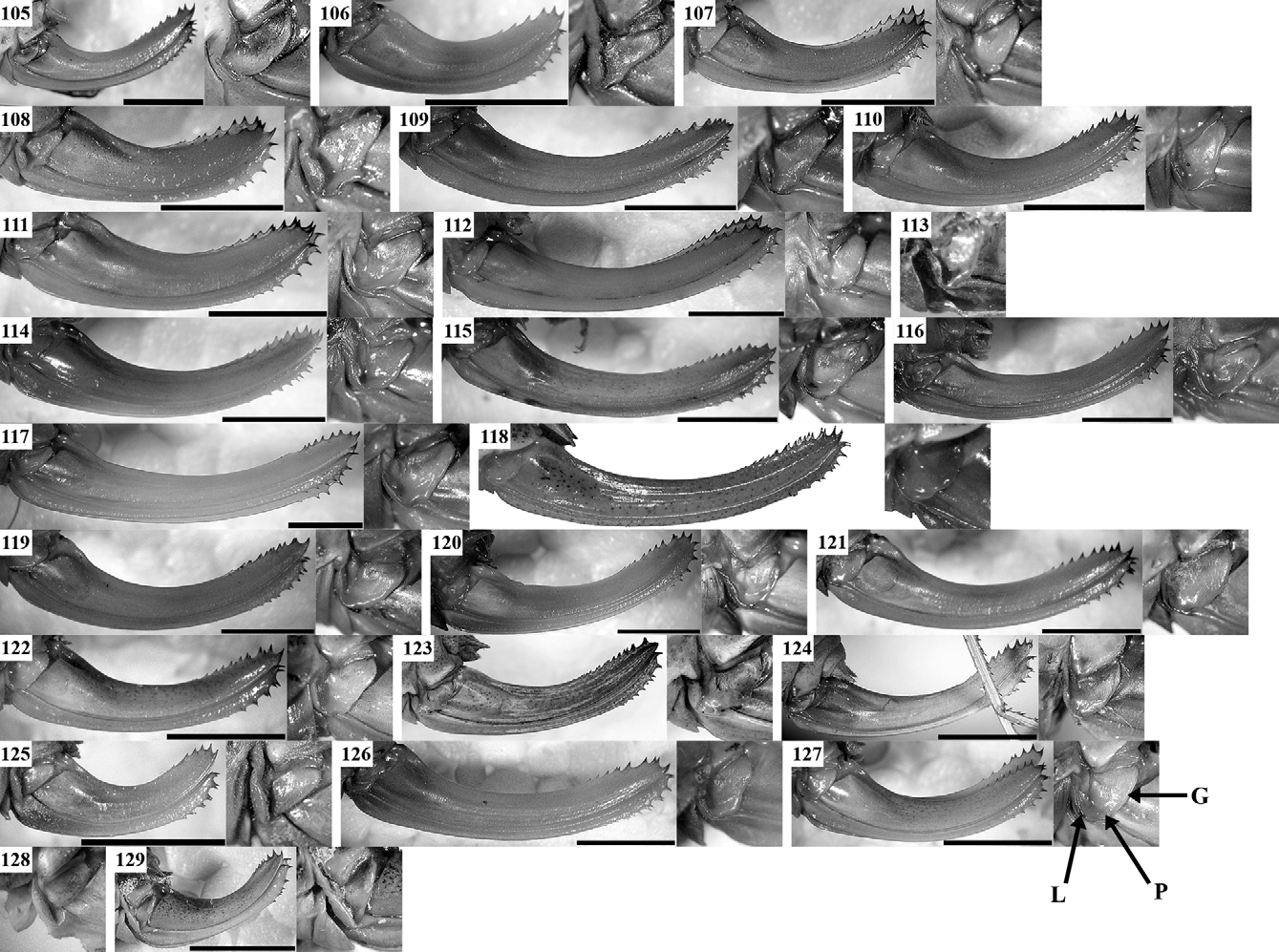
( Figs 1, 2 View FIGURES 1 – 18 , 31, 56 View FIGURES 56 – 79 , 80 View FIGURES 80 – 104 , 105 View FIGURES 105 – 129 , 130, 131 View FIGURES 130 – 134 , 135 View FIGURES 135 – 138 , 191 View FIGURE 191 )
Jaquetia hospodar Saussure : Saussure 1898 (sp.n.).
Isophya medimontana Nedelkov , syn.n.: Nedelkov 1908 (sp.n.). Isophya burmevi Nedelkov : Nedelkov 1908 (sp.n.).
Isophya tschirpanensis Nedelkov : Nedelkov 1908 (sp.n.)
. Isophya hospodar (Saussure) : Kis and Peshev 1967.
Isophya hospodar medimontana Nedelkov : Harz 1969 ( stat.nov.).
Morphological description: see the references above; Harz 1969. Bioacoustics: Heller 1988. Karyotype: Warchałowska-Śliwa et al. 2008.
Synonymy and addition to the description: Nedelkov (1908) described three taxa of Isophya from Bulgaria— I. medimontana , I. burmevi and I. tschirpanensis . Ramme (1951) synonymized the latter with I. burmevi . Kis and Peshev (1967), after redescription and determining the generic belonging of Jaquetia (= Isophya ) hospodar , synonymized I. burmevi and I. tschirpanensis with the latter without mentioning I. medimontana . Harz (1969) provisionally related the latter as a subspecies of I. hospodar pointing that the case could concern synonymy.
After revision of the type series of I. medimontana (see Appendix) and additional material from the region of the Sofia Kettle, Lyulin and Lozenska Planina Mountains, we did not find stable morphological differences between this population and the ones from Dobrogea (the type area) and Southeastern Bulgaria. Some of the specimens from the Sofia Kettle are smaller than the average body size recorded but this may be connected with the microclimate since such small individuals were found also in another region of Bulgaria (Matochina Village in SE Bulgaria). The stridulatory file also does not show differences between populations (compare Figs 130 View FIGURES 130 – 134 A, B and 131A, B). One male from Sofia had file length of 2.2 mm with 68 teeth (52 normally developed and 16 small apical); a stouter male from Eastern Rhodope Mts (SE Bulgaria) showed a file length of 2.65 mm with 73 teeth (57 normal and 16 small); and a male from Gura Dobrogei (SE Romania, the northernmost locality known for this species)—a length of 2.3 mm with 63 teeth. For comparison, Heller (1988) counted 78 teeth in a male from Edirne (European Turkey). The female stridulatory apparatus is shown in Fig. 130 View FIGURES 130 – 134 D. Based on these data we consider I. medimontana as junior subjective synonym of I. hospodar .
Distribution ( Fig. 191 View FIGURE 191 ) and phenology: Endemic species for the Eastern Balkan Peninsula, fragmentary distributed in Southeastern Romania (Dobrogea), Southeastern and Central Bulgaria (westwards reaching the Sofia Kettle), European Turkey and very locally in Northeasternmost Greece. Early species typical for hilly limestone terrains (from the sea level up to 700–800 m) in mesophyte grass associations that become xerophyte in early summer. The species may be found in very low grass associations on pastures. Nymphs—(II)III–V, imago—IV–VI. Notes to the literature distribution data: Peshev (1970b) wrongly refers the locality “Malko Tarnovo” to this species. The case concerns I. pavelii .
2. Isophya rectipennis species group
The group has a number of primitive characters probably derived from these of the I. straubei group. The fastigium is slightly wider, equal (in most taxa) or up to only half as wide as scape (in I. triangularis Brunner von Wattenwyl, 1891). The pronotal disc is not saddle shaped but sometimes the metazone is distinctly wider than the prozone. The length of male tegmona is more or less equal to that of pronotum. The length and width of CuP varies between taxa; CuP and CuA are clearly separated or moderately approximated. The stridulatory file is well developed or partly reduced (in I. rectipennis ) and bears 50–150 teeth. The female stridulatory apparatus has two functional rows of basal spines. The ventral keels of hind femora usually do not possess spines or have single ones. The apical part of male cerci is gradually in- (and sometimes slightly up-) curved, tapered and bearing long pointed or sometimes stout, crest like tooth. The ovipositor is short to moderately elongate, usually less than 2.1 (and always less than 2.3) times longer than pronotum. The basal lateral pit of ovipositor is opened or partly closed; the lamella lacks excision in its distal part. The body colouration is greenish, sometimes with darker or lighter pattern. The lateral margins of pronotum possess reddish stripe only in metazone lying above the light stripe. Melanism is always absent. The song consists of groups or phrases of syllables that may be rapidly repeated in sequences ( I. pavelii , I. nervosa Ramme, 1931 ), simple sequences of syllables ( I. ilkazi Ramme, 1951 —after Heller 1990) or single syllables ( I. thracica , I. cania ). The syllables are short (<100 ms) and decrescending (with a gradual decrease in amplitude after the beginning). The X-chromosome is subacrocentric ( type 1A or 1B according to Warchałowska- Śliwa et al. 2008).
The group after Ünal (2010) includes six species and one additional subspecies. Chobanov (2009a, b) included here also I. thracica and I. cania Karabag, 1975 as intermediate between this group and I. modesta group, though Ünal (2010) places them as relatives of I. schneideri Brunner von Wattenwyl, 1878 and some other taxa. Regarding the morphology, known bioacoustic data, and distribution, both I. thracica and I. cania may show similarities to the I. rectipennis (and partly to I. straubei group in the case of I. thracica as pointed by Karabag 1962) and to I. modesta group (especially in the case of I. cania ) but not to I. schneideri . These characters include for example: (1) form of tegmen—elongated with an obtuse anal corner and narrower than the width of metazone (in I. schneideri the tegmen is broader than the metazone, the anal corner is almost rectangular); (2) tegminal venation in maledistinct separation of CuP and CuA veins (in schneideri— CuP and CuA are closely attached), and in femalemostly with parallel veins, especially laterally, and very weak development of reticulation (in schneideri the venation is reticulate similarly to that of the I. speciosa group—see below); (3) song (see Heller 1988 for I. thracica and Sevgili et al. 2011 for I. cania )—consists of single decrescending syllables followed by an isolated after-click similarly to that of I. modesta group but with short main syllable part as in rectipennis group and in I. bureschi ; on the contrary in schneideri the song is distinctly elaborated, divided into two parts, resulting from incomplete open-and-closing movements similarly to these of the I. speciosa group (Chobanov, unpublished data); (4) lack of melanism while melanism may be partly developed in I. schneideri , which again places the latter closer to the I. speciosa group. The features of I. schneideri also doubt its close relationships with I. sikorai Ramme, 1951 , proposed by Ünal (2010).
Recently, there were two contradictory opinions on the grouping of I. yaraligozi . Though Chobanov (2009b) tentatively relates it to the I. rectipennis group, Ünal (2003, 2010) places it within I. modesta group according to its similarities with I. obtusa mostly concerning the relative length of ovipositor and the cercus shape. However, the mentioned characters are highly convergent in Isophya and cannot be regarded as good markers alone. We relate I. obtusa to another group (for diagnoses of groups see Warchałowska-Śliwa et al. 2008 and the present paperbelow). And though I. yaraligozi is not closely related to I. obtusa the new data on song and morphology of the former support its belonging to the I. modesta group.
Thus, we here consider the I. rectipennis group with 9 taxa distributed in the Eastern Balkan Peninsula and Asia Minor to Lebanon and Syria. We regard the group with three complexes: (1) I. rectipennis complex— I. rectipennis and I. triangularis ; (2) I. pavelii complex— I. ilkazi , I. nervosa , I. pavelii , I. stenocauda stenocauda Ramme, 1951 , I. stenocauda obenbergeri Mařan, 1958 ; (3) I. thracica and I. cania (having acoustic similarities with I. modesta group).





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |