
Isophya obtusa Brunner
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3658.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:C02D1C74-25C0-41DD-B098-62098EB7B62A |
|
DOI |
https://doi.org/10.5281/zenodo.5617396 |
|
persistent identifier |
https://treatment.plazi.org/id/F26F3128-3923-FFBC-B1B0-0CAAFEA29D02 |
|
treatment provided by |
Plazi |
|
scientific name |
Isophya obtusa Brunner |
| status |
|
Isophya obtusa Brunner von Wattenwyl, 1882
( Figs 28, 29 View FIGURES 19 – 30 , 53, 77 View FIGURES 56 – 79 , 102 View FIGURES 80 – 104 , 127 View FIGURES 105 – 129 , 178, 179 View FIGURES 174 – 181 , 188, 189 View FIGURES 182 – 189 , 194 View FIGURE 194 )
Isophya obtusa Brunner von Wattenwyl: Brunner von Wattenwyl 1882 (sp.n.). Isophya pravdini bazyluki Peshev : Peshev 1985 (ssp.n.), syn.n.
Morphological description: see the references above; Bey-Bienko 1954; Harz 1969. Karyotype: Warchałowska- Śliwa et al. 2008.
Synonymy: Peshev (1985) described I. pravdini bazyluki , comparing it with the other subspecies of I. pravdini and thus placing it within the presently regarded I. modesta group. After the revision has been done the other two subspecies of I. pravdini became synonyms of related taxa ( I. plevnensis and I. longicaudata ; see above). Yet, the type series of I. p. bazyluki has not been found. We visited its type locality where, in an isolated spot, an abundant population fitting the description of I. p. bazyluki was discovered. The animals had the same measurements and tegminal structure as given by Peshev (1985) (though the figures in this paper are quite schematic). After comparison of the morphology and song it appeared obvious that we had collected animals of I. obtusa and thus I. pravdini bazyluki is here considered as its junior synonym.
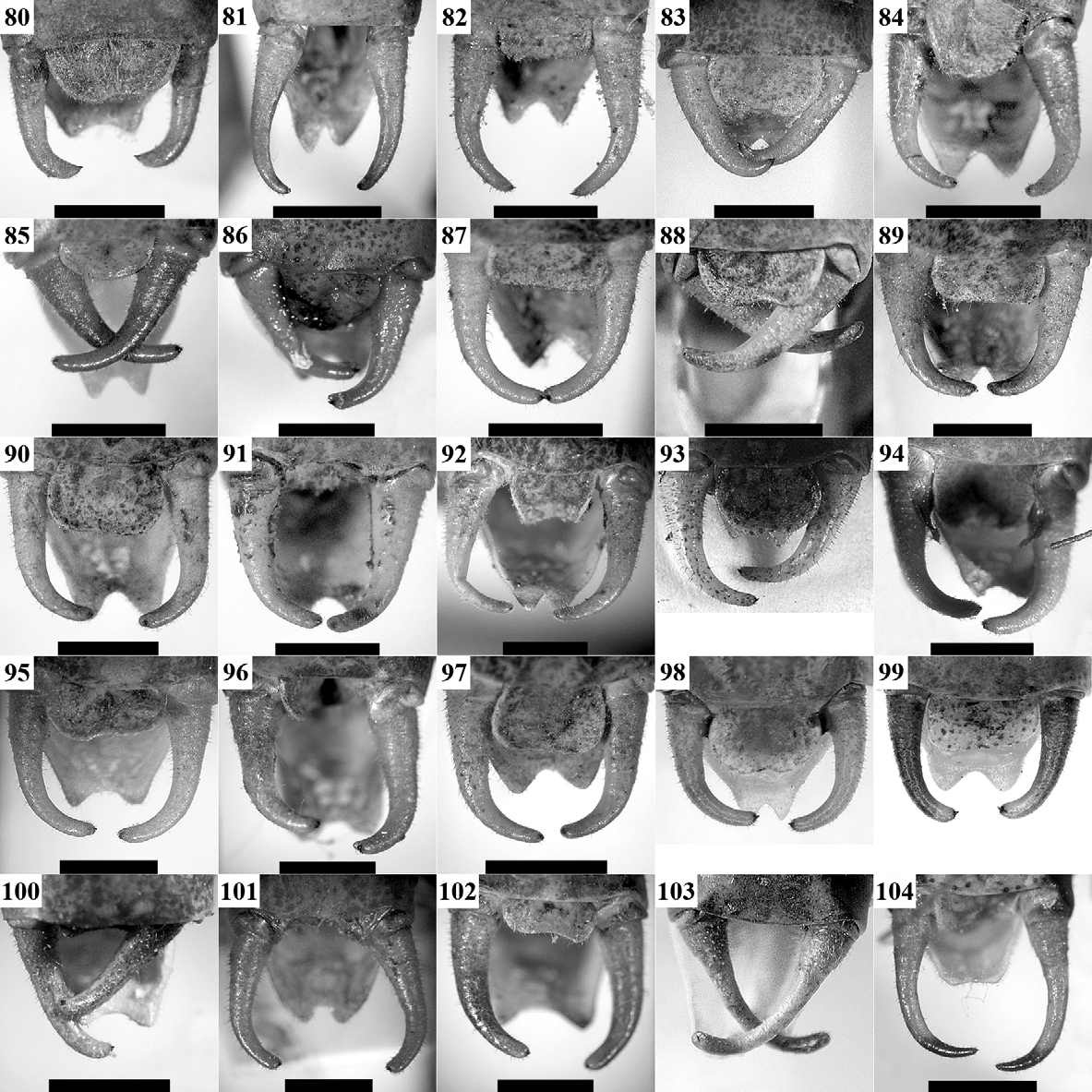
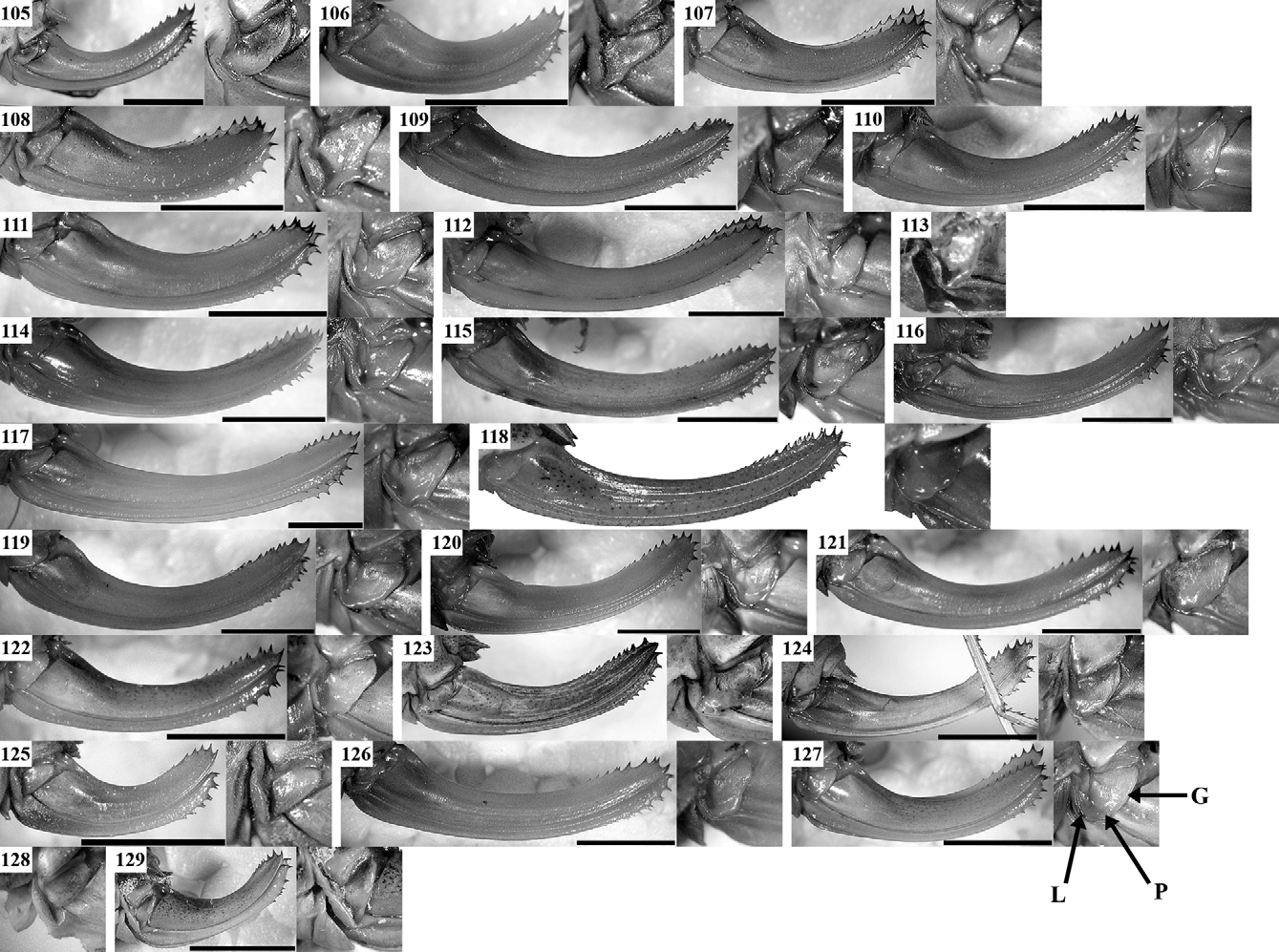
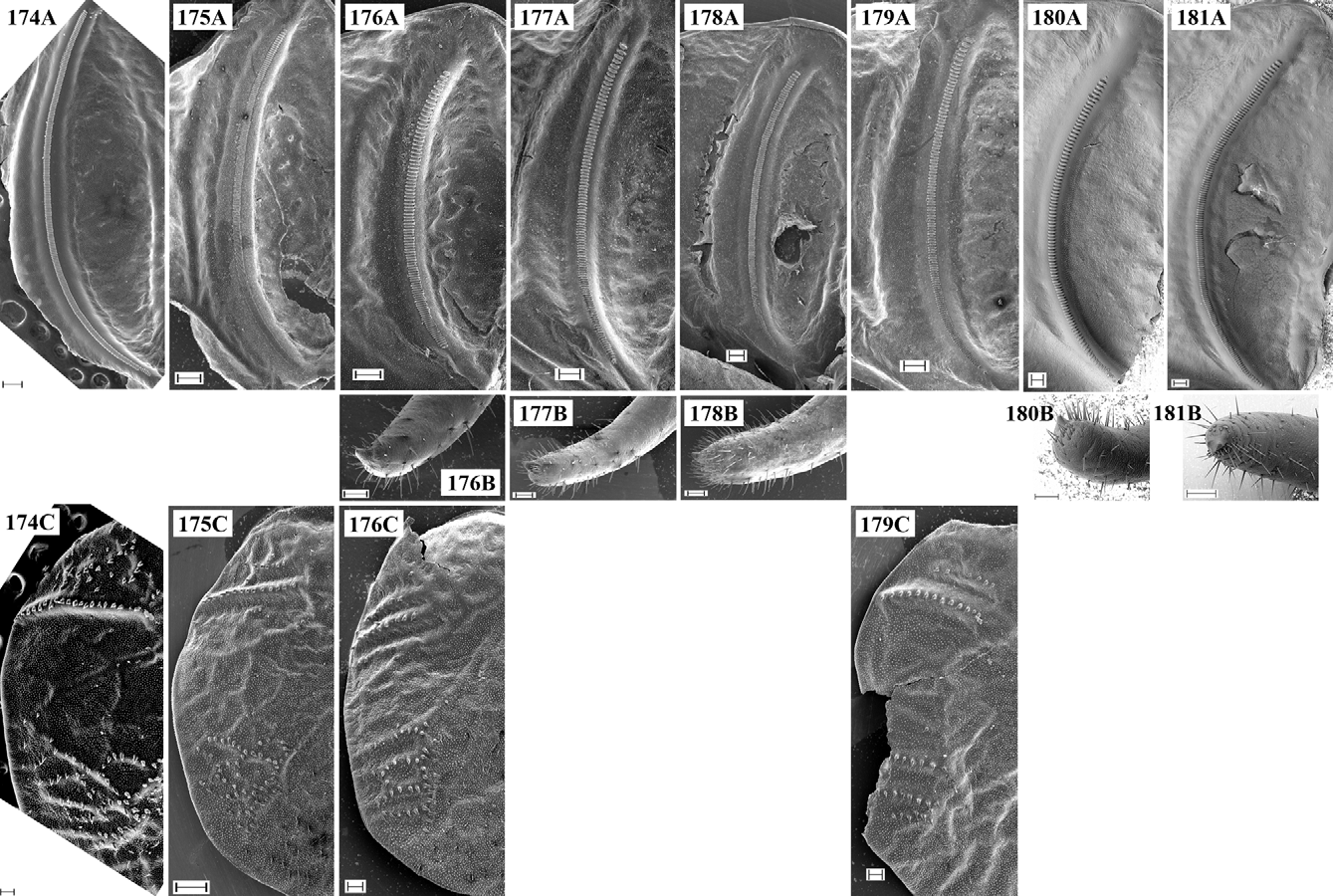
Supplement to the description and diagnosis: The body is moderately stout. Male pronotum ( Figs 28, 29 View FIGURES 19 – 30 , 53) is saddle-shaped, uplifted in prozone and metazone and constricted in its middle; the lateral keels in metazone are sinuate. Male tegmina ( Fig. 53) are equal or slightly longer than pronotum and usually wider than the metazone. The anal edge of left tegmen (at the apical end of CuP) is almost right, frequently protruding over the lateral side of pronotum. CuP is moderately widened and bulged, considerably approximated to CuA, the latter being situated higher than CuP. The stridulatory file ( Figs 178 View FIGURES 174 – 181 A, 179A) has dense teeth and a length from 3.1 mm with 120–125 teeth (specimen from Central Stara Planina Mts) to 3.8 mm with 161 teeth (specimen from Lyulin Mt.); the holotype (“Ilatibor”, ex. coll. Br.v.W., NMW), investigated by Sigfrid Ingrisch, has 137 teeth (according to a label attached to the specimen). Female tegmina ( Figs 77 View FIGURES 56 – 79 , 179 View FIGURES 174 – 181 C) have reticulate venation and are moderately truncate in their apical part. Hind femora have at least one spine on their internal ventral keel. Male cerci ( Fig. 102 View FIGURES 80 – 104 ) are thick, slightly angularly incurved in their apical third (similarly to these of the I. modesta group and I. modestior ) and apically widely obtuse; the apical tooth ( Fig. 178 View FIGURES 174 – 181 B) is wide, short and surrounded by dense hairs. The main colouration is dark green to violet-green with brownish-violet tibiae, tegmina and cerci. The disc of tegmina is brownish to brownish-violet with darker CuP and only the apical part of tegmina may be green. The song ( Figs 188, 189 View FIGURES 182 – 189 ) consists of groups of 2–4 to many (rarely single) gradually decrescending syllables consisting of dense impulses and lasting 100–500 ms. Within the groups the syllables are separated by intervals of 1.5– 4.5 s.
Bioacoustics: The specimens from two separated populations showed some intrapopulation variation and slight interpopulation differentiation of their calling song.
The syllables in the song of specimens from Middle Stara Planina Mountains ( Fig. 188 View FIGURES 182 – 189 ) were frequently separated into a main and an additional part, the latter present or absent within the same recording of an individual. At 25–27°С the main part consisted of 40–70 dense impulses and lasted 100–250 ms (3 33). The impulse period lasted 2.1–3.2 ms at 25°С to 3.5 ms at 27°С. The additional part (when present) followed the main part at 50–170 ms and had 2–6 impulses (after-clicks) and thus the total syllable length became 250–400 ms (3 33). At 28°С the temporal song characteristics of the population from the type locality of I. pravdini bazyluki fit within the above described.
The specimens from Vitosha Mountain showed a song ( Fig. 189 View FIGURES 182 – 189 ) consisting of compact syllables without clearly detached after-clicks, though in some specimens there was a tendence for separating the last few impulses by intervals of 10–80 ms. The syllables lasted 140–420 ms (few 33 at 22 and 25–26°С), whereas the dependence of the length of the syllable of temperature was masked by the high individual variability in the number of impulses (from 30 to 72 counted). The influence of temperature was more clearly recognised in the impulse period, which in the main part at 22°С had an average length of 5.9 ms and at 25–26°С—3.7 ms.
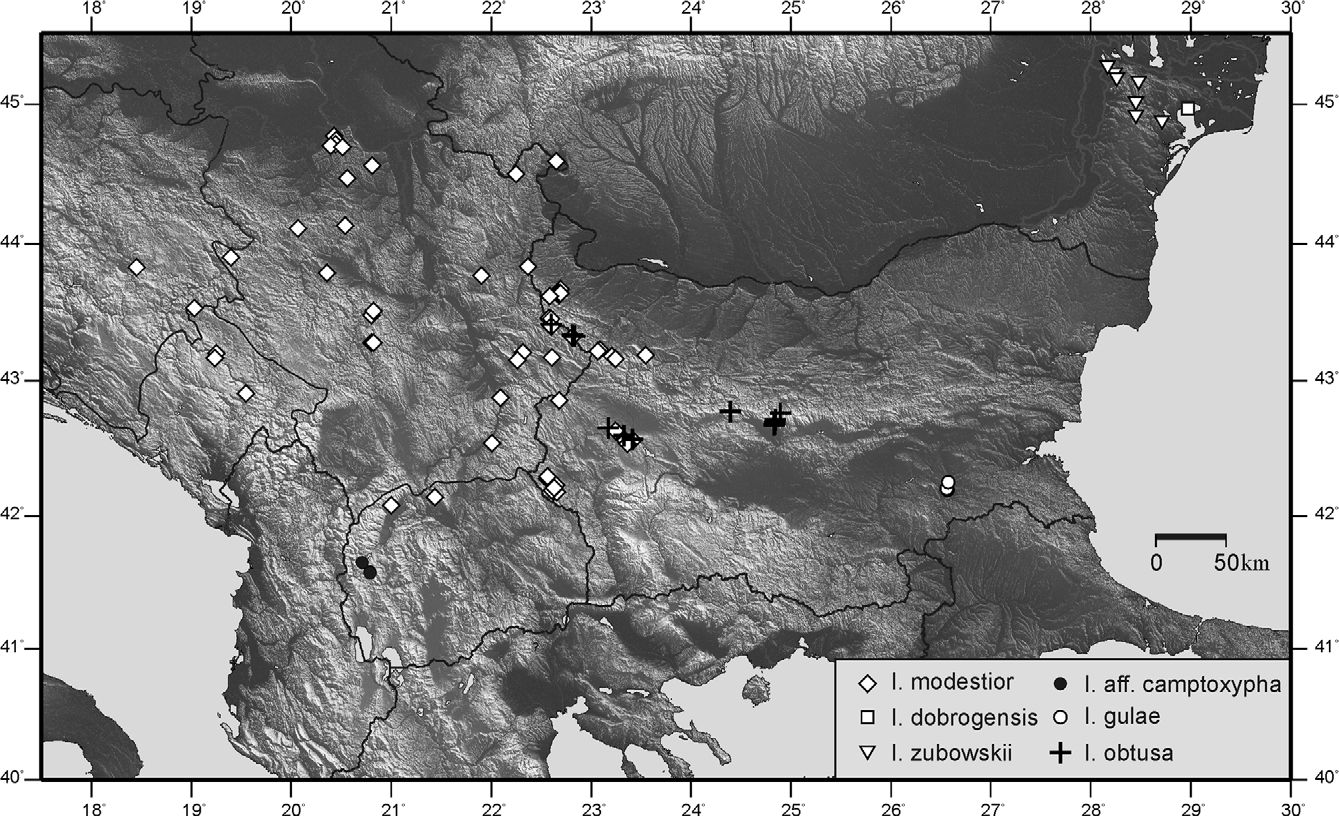
Distribution ( Fig. 194 View FIGURE 194 ) and phenology: The species is fragmentary distributed in the high parts of Middle and Western Stara Planina Mountains and the low- and middle mointain belt of Lyulin, Vitosha and Plana Mountains between 750 and 2150 m alt. (Central and Western Bulgaria and extreme Eastern Serbia). In the lower part of its range it inhabits lush humid plant associations in forests (e.g. observed on Petasites ) while at the upper limit of its distribution may be common in pseudosubalpine mesophyte meadows. Nymphs—IV–VI, imago—VI–XI.
Notes to the literature distribution data: Early records for this species refer to a highly diverse material due to incorrect identification. Data for Sarajevo ( Bosnia and Herzegovina) by Burr (1898) (cited by Us and Matvejev 1967) concern I. clara . The records for Romania (Frey-Gessner 1897; Burr 1899) are also very probably wrong though it is unclear which species is/are concerned in these cases. Data by Ramme (1951) citing Retowskii (1889) for Anatolia (Sinope; Samsun) have been referred to I. amplipennis by Bey-Bienko (1954). The record “Zlatibor” (Brunner von Wattenwyl 1882; Adamoviċ 1975) referred to Zlatibor Mt. in Western Serbia concerns wrong reading the label by Pančiċ where the actual typing is “Ilatibor”. The latter should be referred to a place in Western Stara Planina Mts.
III. Isophya speciosa species group (s.l.)
Warchałowska-Śliwa et al. (2008) outlined the I. schneideri group for a few morphologically similar taxa though they do not give the exact content of the group. Ünal (2010) split this grouping into I. schneideri group (with I. cania , I. hakkarica Karabag, 1962 , I. karabaghi Uvarov, 1940 , I. schneideri , I. sikorai Ramme, 1951 , I. thracica ) and I. amplipennis group (with I. amplipennis , I. artvin Ünal, 2010 , I. gracilis Miram, 1938 , I. hitit Ünal, 2010 , I. redtenbacheri Adelung, 1907 , I. rodsjankoi Bolívar, 1899 , I. savignyi , I. speciosa , I. splendida Naskrecki et Ünal, 1995, I. sureyai Ramme, 1951 ). However, we reconsidered the relationships of few taxa (see above) and related the species I. cania and I. thracica to the I. rectipennis group. I. sikorai should also be excluded from I. schneideri group on account of its morphological position between I. straubei and I. major groups (especially considering the wide fastigium verticis, long tegmina, widely separated CuA and CuP, parallel venation of female tegmina, shape of male subgenital plate and short hind femora with high number of ventral spines); a study of the song is necessary for more precise evaluation of its relationships. Finally, the position of I. karabaghi is also under question since according to the male and female tegminal morphology and male cerci the latter is related to I. bicarinata Karabag, 1957 , a species placed by Ünal in another group ( I. zernovi group).
In I. amplipennis group Ünal (2010) included only the taxa occurring in Turkey while significant number of related species remains unconsidered. Thus, including all morphologically similar species we propose to use the name I. speciosa , the latter being published first.
Isophya speciosa group includes high number of taxa expressing the most specialised characters within this genus. The species have small to moderate body size for the genus and slender appearance. Hind femora are usually shorter than 18 mm. The fastigium is very narrow, less than 1/2 to 1/4 of the width of scapus. Male pronotum is constricted in the middle, widened and raised in pro- and metazone, sometimes clearly saddle-shaped. Male tegmina show smooth transition from a primitive state for the group (mostly in the primitive forms, e.g. I. savignyi , I. rodsjankoi and I. sureyai )—equal or slightly longer and hardly wider than pronotum, slightly bulged, with flat disc and well separated CuA and CuP (or even almost invisible smooth CuA), to a progressive state (in some northeastern and the northwestern forms, e.g. I. speciosa , I. amplipennis )—strongly bulged tegmina, wider and clearly longer than pronotum (up to 2 times longer in I. kalishevskii ), with sharp venation and approximated CuP and CuA (due to a fold in the area between these veins and uplifting the CuA vein). CuP is thin (about as wide as the third antennal limb) and long (usually over 3/4 of the hind margin of metazone). The stridulatory file has 75– 180 teeth (usually about 100); the teeth gradually increase in size towards the apex of file being fine at its base and middle. Female (as well as male) tegmina have clearly developed coarse reticulate venation; their apical part is widely rounded or, rarely, slightly blunt. The lower keels of hind femora do not possess spines. The width and curvature of male cerci vary in different species; the apical spine may be apically or subapically situated, thin or thick, pointed. The ovipositor is very short, shorter than 10 mm (usually less than 9 mm). The pit between the lamella and gonangulum is usually moderately closed by the lamella; the latter may have or have not an excision apically. The body colouration may be green or variable; very typical for the group is developing of melanistic forms with different share of black. The green forms have reddish stripes at the lateral margins of metazone situated above the light band, while in the melanistic forms this stripe is masked by black and red spots. The colouration of male tegmina usually differs from the main body colour and may be variable with yellow, white, red, etc. The calling song consists of highly elaborated single syllables. The song producing movement is generally separated to two consecutive incomplete open-and-closing movements (cf. Heller 1988, 1990) resulting in a syllable consisting of two or more parts. The syllables of the eastern representatives ( I. savignyi , I. gracilis ), corresponding to a more primitive habitus, have short first part and crescending/crescending-decrescending second part, altogether lasting about 500 ms (27.5°С in I. savignyi ; possibly up to one second at lower temperatures) (Heller 1990). In the northwestern taxa ( I. speciosa , I. amplipennis ), showing more progressive morphology, the syllables have three (or even four in I. speciosa ) recognisable parts, produced by distinct slowing down and pausing of each of the two closing movements of the syllable (compare Heller 1988: Abb. 33); altogether the syllable may last three to over five seconds. The X-chromosome is acro- or subacrocentric (type 1B or 2A according to Warchałowska-Śliwa et al. 2008). At least two complexes may be separated within the group, though their exact composition is still ambiguous and needs further investigation (especially of the song structures). The group is most closely related to the I. schneideri group.
I. speciosa group as here considered in a broad sense envelops the highest number of taxa within the genus, distributed from the Caucasus throughout Anatolia to the Balkan Peninsula. About 20 taxa and several groups (e.g. Ünal 2010: partim) are presently considered within the group, most of them occurring in Anatolia, with two species reaching the Balkan Peninsula: I. acuminata , I. amplipennis , I. artvin , I. caspica caspica , I. caspica stshelkanovtzevi , I. gracilis , I. hitit , I. kalishevskii , I. nigrosignata , I. pylnovi , I. redtenbacheri , I. reticulata , I. rizeensis , I. rodsjankoi , I. savignyi , I. speciosa , I. splendida , I. sureyai amazonae , I. sureyai sureyai , I. uludaghensis . Refining the relatioships within the group is at the moment difficult and requires broader phylogenetic study.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |