
Macropelopia nebulosa (Meigen)
|
publication ID |
https://doi.org/10.11646/zootaxa.3852.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:C6CCA5A6-AEC1-4CDF-8F8E-0FB6F157D071 |
|
DOI |
https://doi.org/10.5281/zenodo.6137756 |
|
persistent identifier |
https://treatment.plazi.org/id/F35987B1-FF82-FFD0-FF3C-54A310CEDAE6 |
|
treatment provided by |
Plazi |
|
scientific name |
Macropelopia nebulosa (Meigen) |
| status |
|
Karyotype of Macropelopia nebulosa (Meigen) View in CoL and its relation to Macropelopia paranebulosa (Fittkau)
The chromosome set in M. nebulosa is 2n = 8.The chromosomes are named AB, CD, EF and G on the basis of common band patterns with the chromosome set of M. paranebulosa . ( Makarchenko & Petrova 1988).These authors indicated the chromosome arms of M. paranebulosa by letters S and L. Chromosomes AB, CD, EF of M. nebulosa are metacentric and have large heterochromatin blocks that are particularly noticeable in the middle of the chromosomes. Chromosome G is acrocentric. Very often the chromosomes are conjugated by their centromere heterochromatin forming a structure that looks similar to a chromocenter.
Chromosome AB ( Figure 1 View FIGURE 1 ). The chromosome has a large heterochromatin block near the centromere and is divided into 36 sections.
Arm A has a similar banding pattern to arm S of M. paranebulosa ( Makarchenko & Petrova 1988) . The band patterns 1-2-3- and 17-18-19-20 are similar to those of M. paranebulosa ( Makarchenko & Petrova 1988) . The band patterns between 8–12 are markers of arm A.
Arm B is similar in banding patterns of arm L of M. paranebulosa ( Makarchenko & Petrova 1988) . The banding patterns of arm B are distinguished from that of M. paranebulosa by two steps of homozygous inversions:
39ba -38-37ba-36-35-34-33-32-31-30 -29-28-27-26-25-24-23-22 36-37ab - 38-39ab-35-34-33-32-31-30-29-28-27-26-25-24-23-22
36- 37ab- 38- 39ab-35- 34- 33- 31- 32- 30- 29- 28- 27- 26- 25- 24- 23- 22- 21.
Chromosome CD ( Figure 2 View FIGURE 2 ). The chromosome is divided into 41 sections. The centromere region is characterized by a well defined heterochromatin band and near to this section there is a large heterochromatin block.
Arm C is similar in banding pattern of arm S of M. paranebulosa ( Makarchenko & Petrova 1988) . One step of homozygous inversion distinguishes this arm of M. nebulosa from that of M. paranebulosa .
1-2-3-4-5-6-7-8-9cba- 10-11-12-13-14 -15-16-17-18-22
1-2-3- 4 -5-6-7-17-16- 15-14-13-12 -11-10-9abc- 8-18-22.
Arm D is similar in banding patterns to the arm L of M. paranebulosa ( Makarchenko & Petrova 1988) . Three steps of homozygous inversions distinguished arm D of M. nebulosa from that of M. paranebulosa .
41-40-39-38-37-36-35-34-33-32ab-31-30ab-29-28-27-26-25-24-23-22 41-26-27-28-29-30ba-31-32ba-33-34-35-36-37-38-39-40-25-24-23-22 41-26-27-28-29-30ba-32ab-31-33-34-35-36-37-38-39-40-25-24-23-22
41-26-27-28-29-30ba-32ab-31-37-36-35-34-33-38-39-40-25-24-23-22
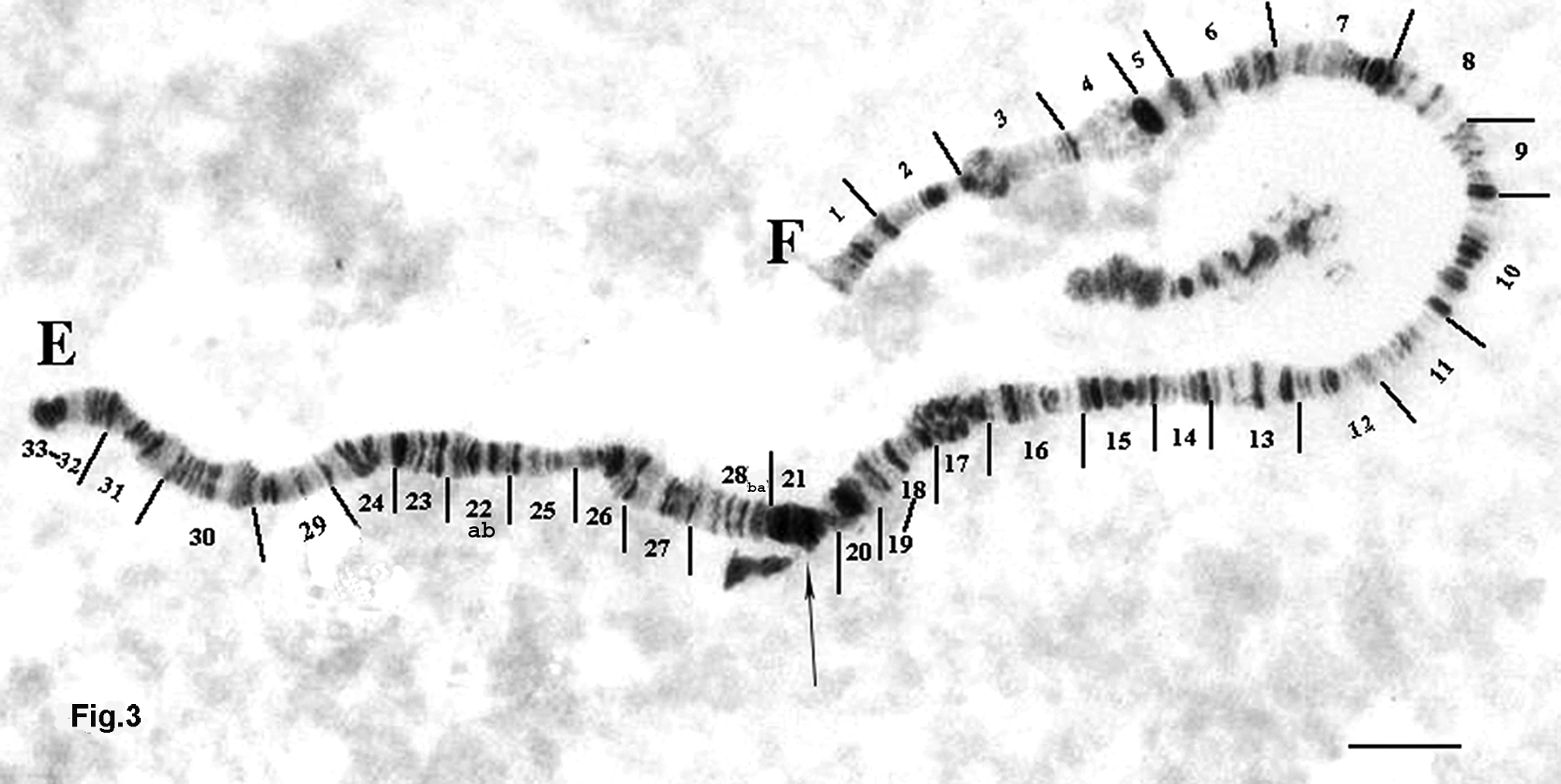
Chromosome EF ( Figure 3 View FIGURE 3 ) is divided into 33 sections.
Arm E is similar to banding patterns of arm S of M. paranebulosa ( Makarchenko & Petrova 1988) . Two steps of homozygous inversion distinguishes arm E of M. paranebulosa from that of M. nebulosa .
33-32-31-30-29-28ab-27-26-25-24-23-22ab-21- 33-32-31-30-29-22ba-23-24-25-26-27-28ba- 21 M. nebulosa
33-32-31-30 -29-24-23-22ab-25-26-27- 28ba- 21 The banding patterns 23-22ab-25-26-27-28ba are markers of the arm E.
Arm F is similar to banding patterns L of M. paranebulosa ( Makarchenko & Petrova 1988) . The band patterns 1-2-3-4-5- of arm F of M. nebulosa are very similar to those of M. paranebulosa . Similarity was also found between band patterns 20-19-18 of both species. From section 7 to section 18 the band patterns of M. nebulosa are indicated conditionally because the equivalent bands of M. paranebulosa are not clear and hence comparative analysis is not possible.
1-2-3-4-5-6-7-8-9- 10 -11-12-13-14-15-16-17-18-19-20
Banding patterns of the specimens from the UK localities have a homozygous inversion in section 4–13. The band patterns 12-13-14-15 -16-17- of arm F are markers of this arm.
Arm G ( Figure 4 View FIGURE 4 ) is divided into 4 sections.
This arm of M. nebulosa is distinguished from that of M. paranebulosa by simple homozygous inversions.
M. paranebulosa NOR1-2abc-3ab-4
NOR1 -2cba-3ab-4.
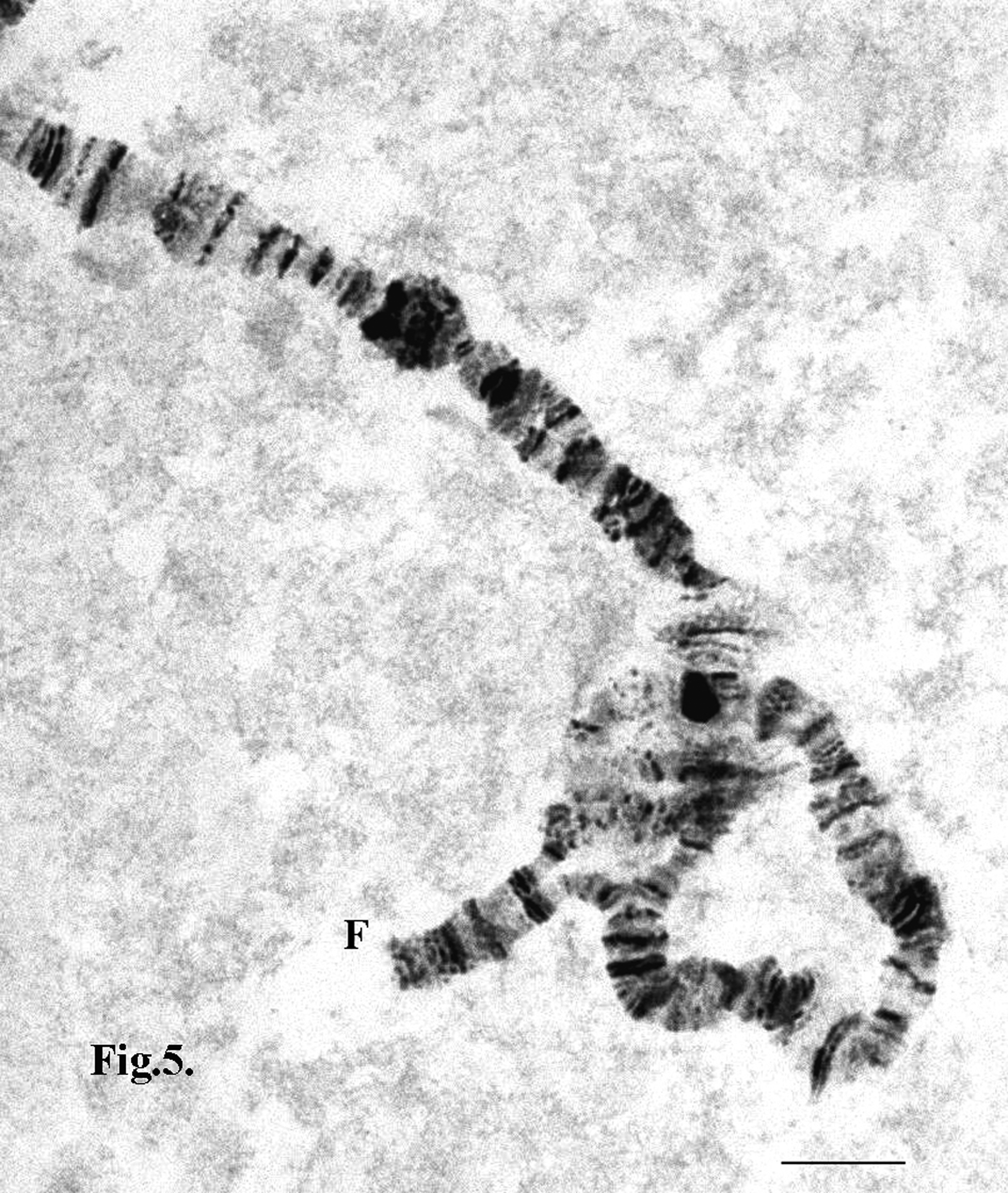
In nine specimens from Bulgaria heterozygous inversions in two types were found: inherited and somatic aberrations. Inherited affected all cells of the specimens. The complex inherited heterozygous inversions were detected in arms A, B, C, D and F ( Figure 5 View FIGURE 5 ). Somatic aberrations affected few cells only. These types of rearrangements w e r e established in single cells and affected arms A, B, D, F and E from species in Bulgaria. In the specimens collected from the UK no homokaryotype was found ( Figures 6 View FIGURE 6 a ab). In all specimens both inherited and somatic rearrangements were detected. Complex inherited heterozygous inversions were found in all chromosome arms, while somatic rearrangements were located mainly near to the centromere regions of chromosomes AB and CD, as well as in the middle of arms A, B, D. Some heterozygous inversions in t h e somatic state were found towards the telomere of arm E. In two specimens from the Southern Afon Goch at City Dulas locality the deletions of arm G in somatic appearance were detected.
Despite an improvement in water quality due to remediation measures earlier this century (see Dean et al. 2013) water and sediment analysis from the UK sites all showed a high degree of contamination by the metals copper, zinc, iron and manganese. Concentrations of all metals exceeded standards and guidelines for both sediments and water (e.g. Abdullah & Royle 1972; Warren 1981; MacDonald et al. 2000; Irish EPA 2001). However, the water at all sites is slightly acid to near-neutral ( pH 5.8–7.3) which allows colonization by M. nebulosa . In Bulgaria the species was found in a locality characterized by municipal contamination only. The somatic rearrangements described here can thus be used as biomarkers for contaminants in water basins ( Michailova et al. 2012), including trace metals and organic pollutants.
Both M. paranebulosa View in CoL and M. nebulosa View in CoL had 2n = 8, but differed in their banding patterns of the salivary gland chromosomes. These differences can be explained by chromosome rearrangements (fixed homozygous inversions) that have affected the chromosome morphology but not the chromosome set of both species. The divergence of both species passed on these types of aberrations which can be sufficient to act as post-mating reproductive barriers ( King 1993). The application of cytogenetic approach for Chironomid identification has been recognized as a useful tool to resolve the problems of species identification in the Tanypodinae View in CoL subfamily. M. paranebulosa View in CoL and M.nebulosa View in CoL are sibling species ( Makarchenko & Petrova 1988). In external morphology of larvae the both species cannot be distinguished. However, M. nebulosa View in CoL larvae have species-specific cytogenetic markers by which the species can be identified. The cytogenetic method once again has proven useful in identifying Chironomid at the larva stage despite the salivary gland chromosomes of M.nebulosa View in CoL being very long and spread with difficulties. This is reliable and quick approach for routine identification of species of this and other chironomid genera. The chromosome map of M. nebulosa View in CoL presented in this work can now be used successfully in genotoxicological studies.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Tanypodinae |
|
Genus |