
Acetes, H. MILNE EDWARDS, 1830
|
publication ID |
https://doi.org/10.1111/zoj.12371 |
|
persistent identifier |
https://treatment.plazi.org/id/F51387D8-1623-9348-FB9F-E156B7E8F9A9 |
|
treatment provided by |
Marcus |
|
scientific name |
Acetes |
| status |
|
AND ACETES
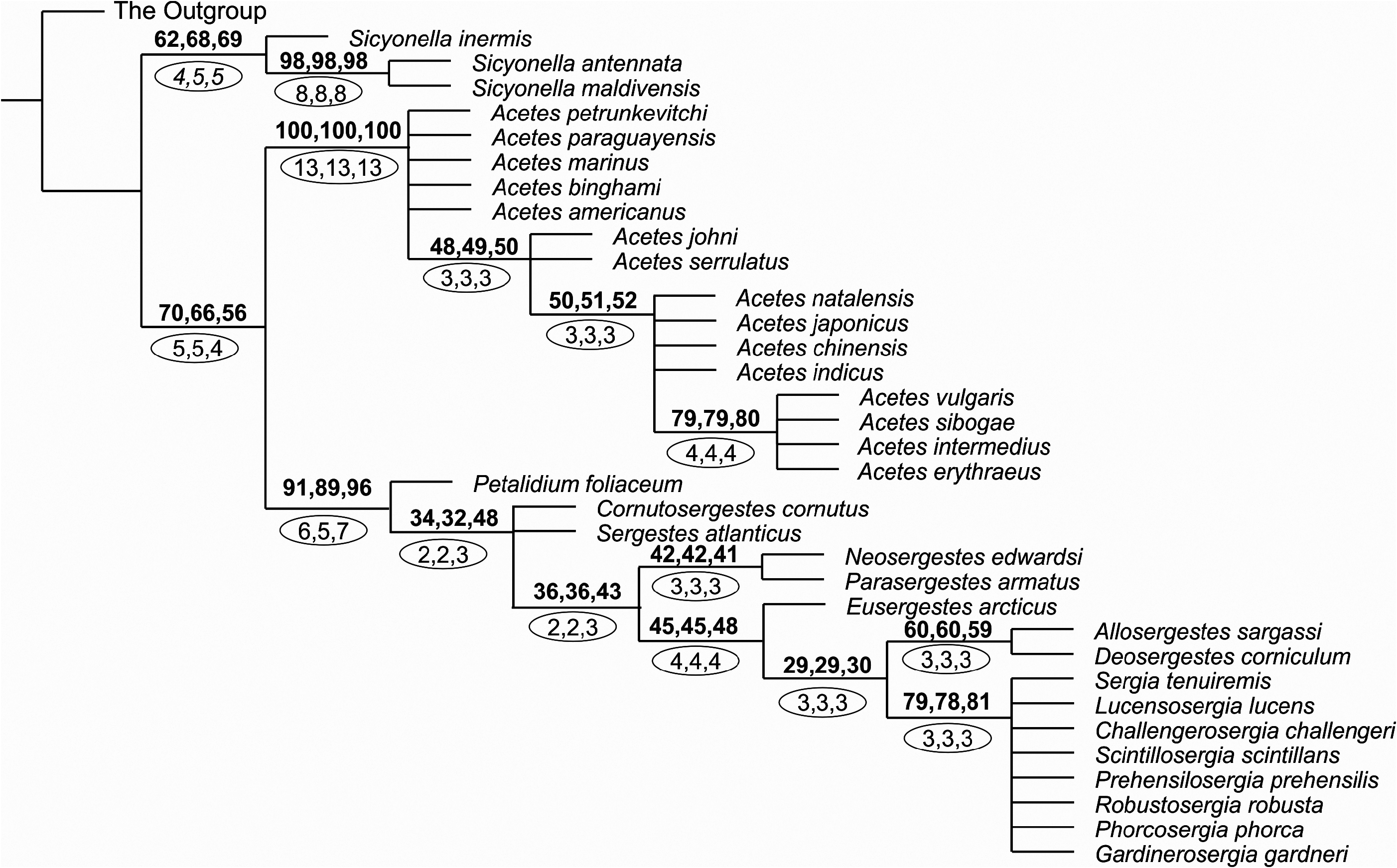
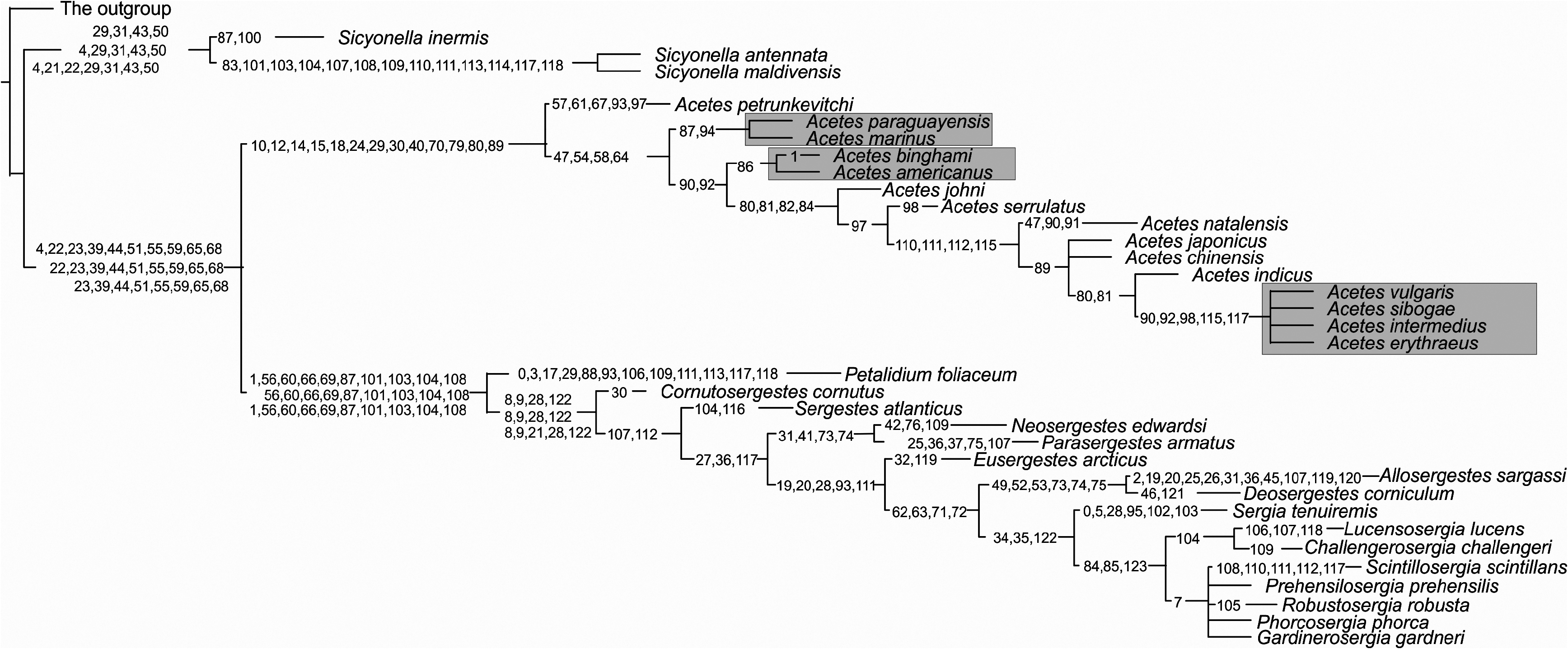
Morphological analyses show that Sicyonella differs most clearly from the rest of Sergestidae in characters that may be associated with its occurrence at the benthopelagic shelf. These characteristics may provide additional protection, manoeuvrability, and feeding opportunities near the water–bottom interface ( Vereshchaka, 1990, 1995). Some of these characters are synapomorphic ( Fig. 12 View Figure 12 ): two endites at the maxillulae (character 29 – see Appendix 2), enlarged third maxillipeds (31), and welldeveloped chelae of second and third pereopods (43, 50). Other characters (two dorsal teeth on the rostrum, pterygostomial tooth, four moveable lateral spines on the telson, a complete set of segments in fourth and fifth pereopods) may have been inherited from a common penaeid ancestor.
Most other characters within Sicyonella are associated with mating (male petasma and coupling struc- tures). Hence, the species S. inermis is characterized by the presence of fine scales opposite the tubercle in the clasping organ (87) and by the presence of squamose hooks (100) the entire capitulum of the petasma ( Fig. 12 View Figure 12 ). In addition, another clade, S. antennata + S. maldivensis , is supported by a set of synapomorphies related to mating and coupling ( Fig. 12 View Figure 12 ): claw-like setae at the clasping organs opposite the tubercle (83), a divided capitulum of the petasma armed with squamose hooks and pincers (101, 104), well-developed lobi armatus, connectens, and terminalis (104, 107, 108–109), well-developed, elongated, twice-branched processus ventralis (110,111, 113, 114, 117, 118). The claw-like setae of the clasping organs may have a holding function. The petasmas of S. antennata and S. maldivensis are probably the most elaborate within Dendrobranchiata, with all known processi and lobi and the branched procesuss ventralis being present at the same time.
The clade Acetes is supported by synapomorphies related to the reduction of the branches (10, 12, 14, 15, 18), of the mouthparts (29–30), of the first chelae (40), and of the pereopods (adapted for swimming) (70). These characters, along with the general reduction in size and body compression, are probably adaptations to estuarine and freshwater shallow habitats, and to the fast reproductive cycles observed in the genus ( Omori, 1975). These habitats have permanently high concentrations of oxygen, provide no possibility for extensive vertical migrations (too shallow), are productive, and in general favour short life cycles; some species have two generations in a season ( Omori, 1975). Yet another set of synapomorphies is related to the presumed coupling procedure: the elongated third antennular segment (24), the presence of two clasping tubercles (79–80), and the strong distal tooth in the clasping organ (89).
All species of Acetes , except A. petrunkevitchi , show further reduction and loss of structures related to movement (fourth and fifth pereopods – Fig. 12 View Figure 12 ) that may be further adaptations to shallow habitats where significant vertical migrations are impossible. Within Acetes , the phylogeny is based only on sexual characters, e.g. the clasping organ and the petasma ( Fig. 12 View Figure 12 ).
Omori (1975: fig. 3) manually depictured six clades of Acetes : (1) A. marinus + A. paraguayensis , (2) A. erythraeus + A. intermedius + A. sibogae + A. vulgaris , (3) A. indicus , (4) A. serrulatus + A. johni + natalensis , (5) A. japonicus + A. chinensis , and (6) A. americanus + A. binghami
However, our analysis supports only three of Omori’s clades ( Fig. 12 View Figure 12 ):
1. The clade A. marinus + A. paraguayensis , which is not statistically robust but supported by the presence of fine scales on the clasping organ opposite the tubercle (87) and by the absence/strong reduction of the capitulum of the petasma (94).
2. The clade A. americanus + A. binghami , which is also not statistically robust but supported by the presence of specialized serrated setae with longitudinal ribs in the clasping organ (86).
3. The terminal clade A. erythraeus + A. intermedius + A. sibogae + A. vulgaris , which is statistically robust ( Fig. 11 View Figure 11 ) and supported by the following synapomorphies of the petasma: the presence of well-developed pars astringens (90, 92), additional enlarged claws on the capitulum (98), and an elongated processus ventralis (115, 117).
Most of the morphological variation within Acetes is seen in sexual characters, but because of substantial homoplasy, the phylogeny within the genus is hard to resolve. Like Sicyonella , Acetes is supported by synapomorphies that may be regarded as adaptations to estuarine habitats. Once adapted to such a habitat, the subsequent speciation within both genera has seemingly only been related to mating mechanisms (clasping and copulation).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.