
Dysmicoccus kunaw Tanaka, 2022
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5168.3.3 |
|
DOI |
https://doi.org/10.5281/zenodo.6907996 |
|
persistent identifier |
https://treatment.plazi.org/id/F57C87F7-E97B-FF97-FF03-FA1EFEDCFAD2 |
|
treatment provided by |
Plazi |
|
scientific name |
Dysmicoccus kunaw Tanaka |
| status |
sp. nov. |
Dysmicoccus kunaw Tanaka sp. nov.
(Japanese common name: Tanpopo-kona-kaigaramushi)
Material examined. Holotype: JAPAN, Hokkaido, / Kamikawa-gun, / Pippu-cho, / Kamikawa Agricultural / Experiment Station , / on Taraxacum officinale , / 9.vii.2020, / coll. D. Sasaki; adult female mounted singly ( ELKU) . Paratypes: same data as for holotype; 9 adult females mounted singly (4 ELKU, 5 EUMJ) . Paratype used for molecular phylogenetic analysis: same data as for holotype ; 1 adult female (1 ELKU, Genbank Accession No. ON533755 View Materials for COI, ON527952 View Materials for 18S, ON527954 View Materials for 28S D2) .
Description (n = 11). Live adult female: Feeding on the roots of the host plant and secreting white powdery wax on all body surfaces ( Fig. 2 View FIGURE 2 ); the mealybugs are attended by the ant Lasius flavus Fabricius ( Hymenoptera : Formicidae ).
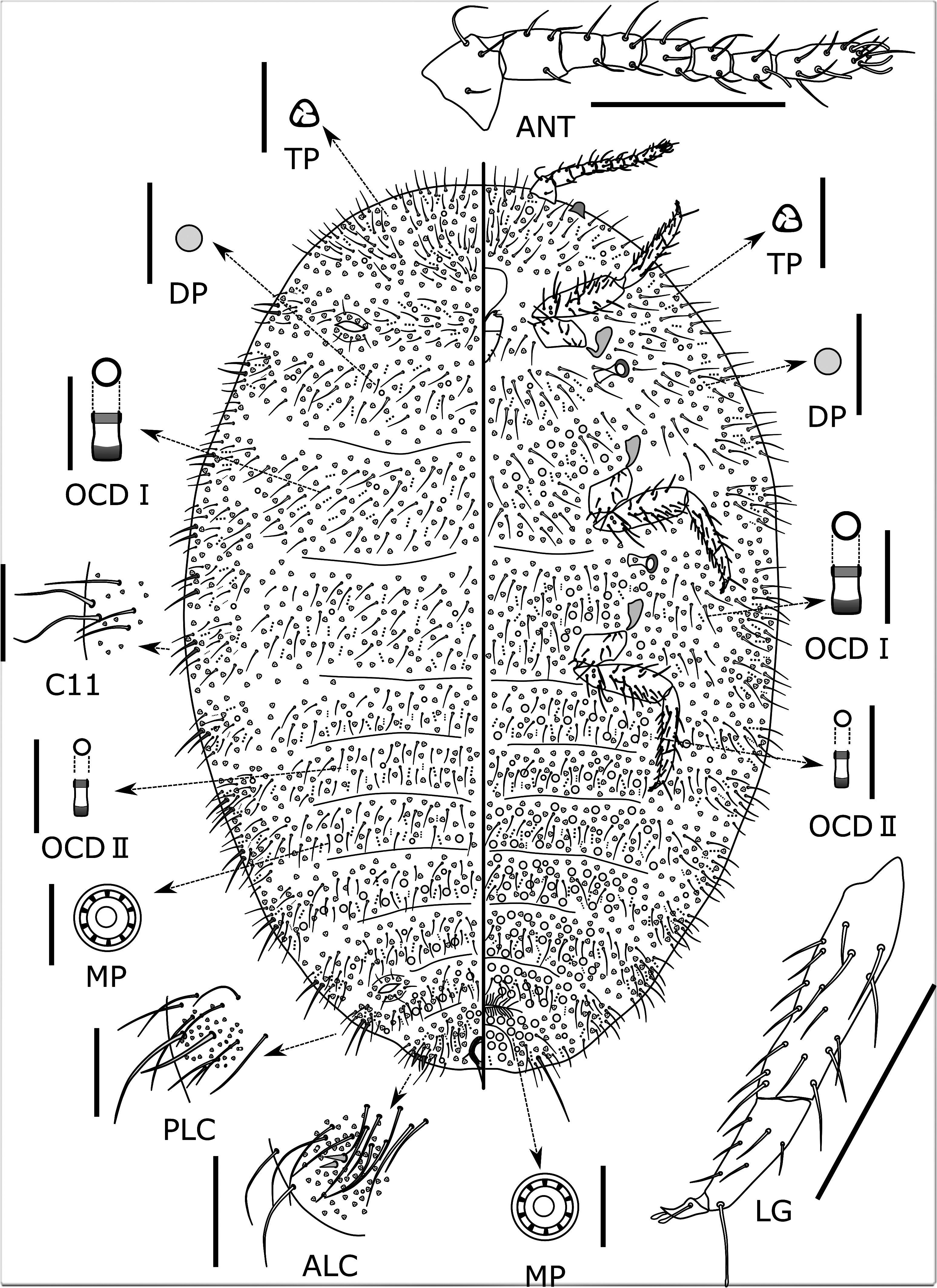
Slide-mounted adult female ( Fig. 3 View FIGURE 3 ): Body elongate oval, 3.1 (2.4–3.1) mm long and 2.0 (1.5–2.2) mm wide; derm membranous; segmentation not well-developed. Anal lobes clearly evident but not prominent, dorsal and ventral surfaces of each lobe without sclerotised areas and ventral surface with long apical seta, 160–225 (150– 235) µm long. Antenna 439–444 (375–447) µm long, with 7 or 8 segments (usually 8) and many flagellate setae; subapical segment with 1 (0 or 1) fleshy seta and apical segment with 3 (2–4) fleshy setae. Eyes present on margin, not associated with discoidal pores. Legs well-developed, with many flagellate setae; hind trochanter + femur 333–344 (295–348) µm long; hind tibia + tarsus 330–334 (295–340) µm long; claw 40–41 (36–43) µm long without a denticle. Ratio of lengths of hind tibia + tarsus: trochanter + femur about 1: 1.0 (0.9–1.0); ratio of lengths of hind tibia to tarsus 1: 1.8–2.0 (1.7–2.1). Single or paired setose tarsal digitules present, subequal in length to minutely knobbed claw digitules. Hind legs without translucent pores. Labium 120 (120–136) µm long, shorter than clypeus. Circulus absent. Ostioles present, each with inner edges of lips weakly sclerotised; anterior ostioles each with a total for both lips of 22–28 (12–28) trilocular pores and 5 or 6 (1–6) setae; each posterior ostiole with a total for both lips of 42 or 43 (16–47) trilocular pores and 1 or 2 (0–4) setae. Anal ring 90 (78–101) µm wide, with 2 rows of cells, bearing 7 (6 or 7, usually 6) setae, each seta 110–161 (60–170) µm long. Cerarii numbering 15–17 (14–17) pairs. Anal lobe cerarii mostly each containing 2 slender conical cerarian setae, each seta 16–20 (15–20) µm long and about 6–7 (6–8) µm wide at base, also 10–13 (7–16) auxiliary setae and a concentration of trilocular pores. Penultimate cerarii each containing 2 long, stout flagellate cerarian setae and many auxiliary setae. Cerarii situated further forward generally each with 1–3 stout long, flagellate cerarian setae and auxiliary setae; boundary of each cerarius on anterior thoracic segments and head obscure and sometimes difficult to distinguish.
Dorsum. Setae long and flagellate, each 49–132 (40–162) µm long, distributed segmentally. Trilocular pores, each 3–4 µm wide, evenly distributed. Oral collar tubular ducts of 2 sizes: (i) large-type ducts, each about 3–4 µm in diameter, mostly each same width as a trilocular pore, present on all segments of dorsum, forming transverse bands across most segments; and (ii) small-type ducts, each about 1–2 μm in diameter, relatively sparse on all segments, intermixed with large-type ducts. Discoidal pores, each 2–3 µm wide, sparsely distributed. Multilocular disc pores each 7–9 (7–10) µm wide, present on medial area of abdominal segments as follows: segment I, 0 (0); II, 0 (0 or 1); III, 1 (0 or 1); IV, 5 (0–6); V, 0 (0–14); VI, 2 (2–10); VII, 7 (1–11); and VIII, 0 (0 or 1).
Venter. Setae long and flagellate, each 64–160 (25–194) µm long, distributed segmentally, longest on medial area of head or posterior abdominal segments. Multilocular disc pores, each 7–9 (7–11) µm wide, present on medial area of all segments of venter between head and abdominal segment IX. Trilocular pores, same width as on dorsum, evenly distributed. Oral collar tubular ducts of 2 sizes: (i) large-type ducts, each about 3–4 µm in diameter, mostly same width as trilocular pores, present on all segments of venter, forming transverse bands across most segments; and (ii) small-type ducts, each about 1–2 μm in diameter, relatively sparse between mesothorax and abdominal segments VII, intermixed with large-type ducts. Discoidal pores, same width as on dorsum, sparsely present.
Host plants. Roots of Taraxacum officinale (Asteraceae) .
Remarks. Dysmicoccus kunaw resembles D. trispinosus ( Hall 1923) described from Egypt, and D. furcillosus Williams 2004 described from India, in having a slightly rounded body, long flagellate body setae, and long flagellate cerarian setae. However, D. kunaw differs from them as follows (contrasting character states in D. trispinosus and D. furcillosus in parentheses): (i) each anal lobe cerarius with two conical cerarian setae (anal lobe cerarian setae not conical, but long and stout setose); (ii) abdominal segments of dorsum with a considerable number of multilocular pores, also on head and thoracic segments of venter (multilocular disc pores absent from dorsum, and head and thoracic segments of venter); and (iii) ventral oral collar tubular ducts of two types (only one type of ventral duct present in D. trispinosus , and three types in D. furcillosus ).
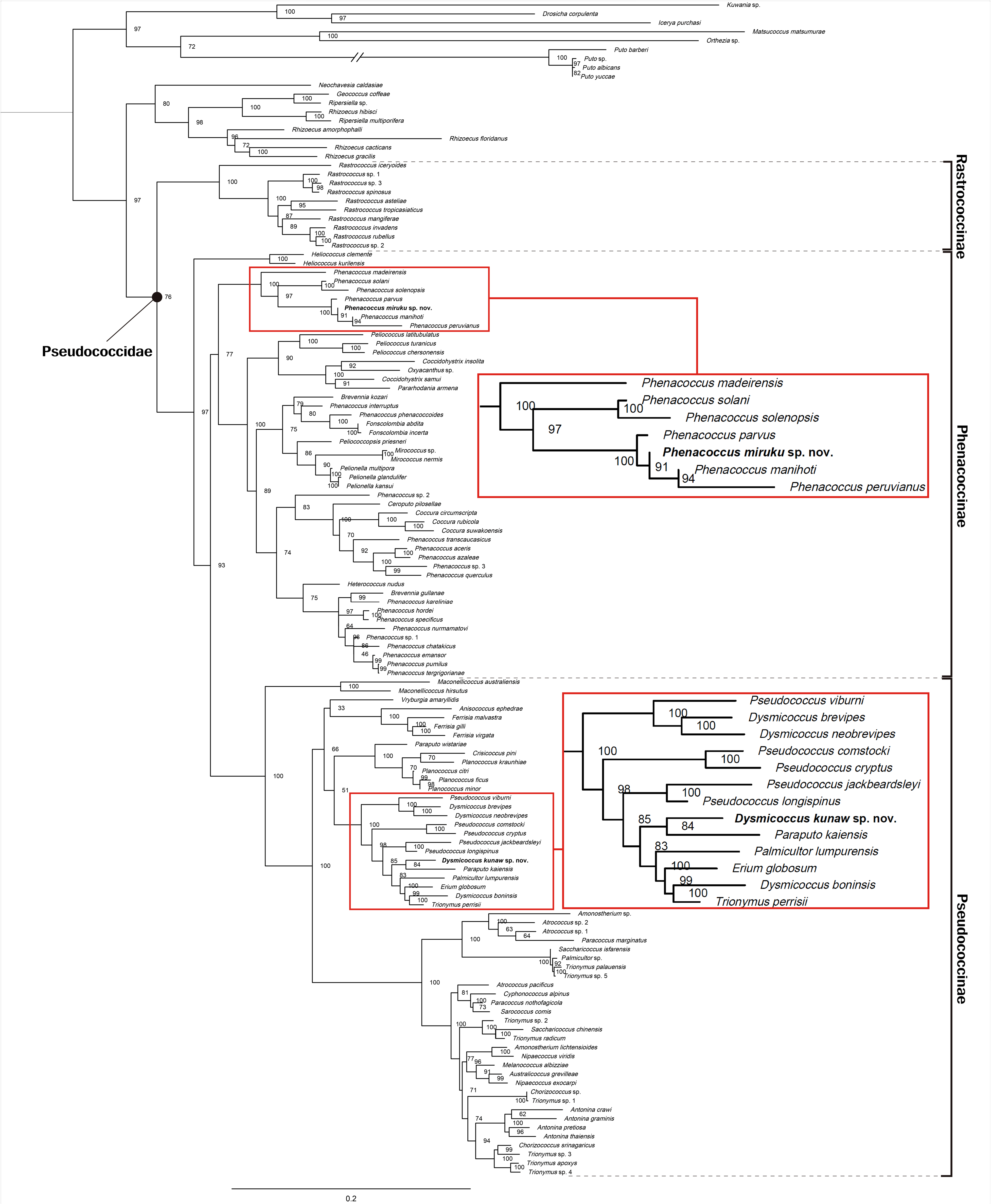
The molecular phylogenetic analysis placed D. kunaw within the clade of the subfamily Pseudococcinae ( Fig. 1 View FIGURE 1 ). The new species formed a clade with Paraputo kaiensis ( Kanda 1932) , although this was not well supported (UFBoot = 84) and showed relatively long branch lengths. This clade was sister to a clade including species of Dysmicoccus , Erium , Palmicultor and Trionymus . A clade with Pseudococcus cryptus Hempel 1918 and P. jackbeardsleyi Gimpel & Miller 1996 was placed outside the clade containing D. kunaw .
The molecular analysis does not support the generic designation of the new species in this study, which is based on adult female morphological characteristics. The analysis also indicated that D. kunaw was not related to the type species of Dysmicoccus , the neotropical species D. brevipes (Cockerell 1893) . Dysmicoccus is known to be a polyphyletic group ( Choi & Lee 2022). Although the genus Dysmicoccus needs revision, we tentatively place D. kunaw in Dysmicoccus based on the current morphological classification.
Relationship with ants. The type series of D. kunaw was tended by the ant Lasius flavus Fabricius 1782 ( Hymenoptera : Formicidae ) and was found inside ant-made carton shelters. Dysmicoccus kunaw may have a close relationship with this and other ant species belonging to the genus Lasius .
Etymology. The specific epithet “ kunaw ” is an Ainu noun that refers to a dandelion, a forked-stem adonis, or a goddess of mist in Ainu mythology who was transformed into a flower by the curse of her husband, Hoinu, the marten god. The Ainu are the local indigenous people of Hokkaido Island. The epithet is used as a noun in apposition.

| EUMJ |
Ehime University |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |