
Megophrys kalimantanensis, Munir & Hamidy & Matsui & Iskandar & Sidik & Shimada, 2019
|
publication ID |
https://doi.org/10.11646/zootaxa.4679.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:5820FB68-81D5-4D5A-8E0E-8083BD06D21E |
|
persistent identifier |
https://treatment.plazi.org/id/F87DDD1A-BF65-3112-FF6D-FF49FDAFDAA6 |
|
treatment provided by |
Plazi |
|
scientific name |
Megophrys kalimantanensis |
| status |
sp. nov. |
Megophrys kalimantanensis sp. nov.
( Figs. 4 View FIGURE 4 , 5 View FIGURE 5 , 6 View FIGURE 6 , 7 View FIGURE 7 , 8 View FIGURE 8 & 10 View FIGURE 10 )
Megophrys nasuta View in CoL (part): Frost et al. 2006, p. 310; Hamidy et al. 2012, p. 26; Matsui et al. 2017a, p. 3.
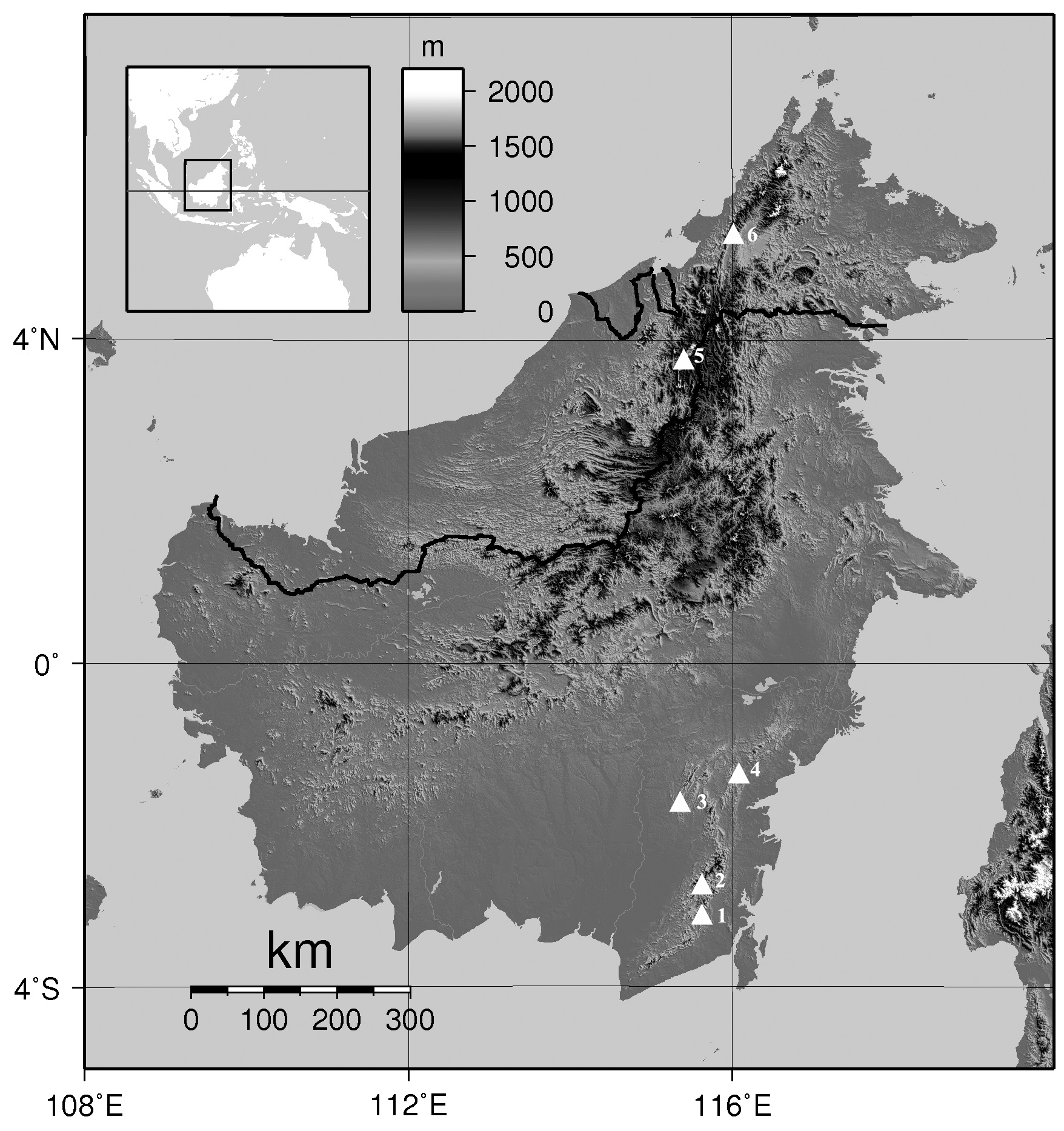
Holotype ( Figs. 4 View FIGURE 4 , 5 View FIGURE 5 A–E). MZB. Amph 31765, an adult male from the western slope of Meratus Mountain Range , Jumantir Camp Site , (Desa) Village of Juhu , (Kecamatan) District of Batang Alai Timur , (Kabupaten) Regency of Hulu Sungai Tengah, in the province of Kalimantan Timur, Borneo, Indonesia (- 2.684389° S, 115.615167° E, elevation 1000 m a.s.l.), collected by Tomohiko Shimada, Ahmad Nabil Faturahman and Huda Wiradarma on 17 September 2018 at 18.00 h ( Fig. 1 View FIGURE 1 ). GoogleMaps
Paratypes. MZB. Amph 31766 and 31767 two adult males collected together with the holotype ; MZB. Amph 25054, 25055 and 25056, three adult males collected from the western slope of Meratus Mountain Range, Emil Baru Village , Mantewe District , Tanah Bumbu Regency , Kalimantan Timur (- 3.079500° S, 115.581028° E, elevation 943 m a.s.l.) by Irvan Sidik on April 2008 GoogleMaps ; MZB. Amph 25436, a subadult female, collected on February 2009 from the Meratus Mountain Range, Paniti Ranggang, Juhu Village , Batang Alai Timur District , Hulu Sungai Tengah Regency , Kalimantan Selatan (- 2.643242° S, 115.594647° E, elevation 542 m a.s.l.) GoogleMaps ; MZB. Amph 25728, an adult female, collected on May 2014 from Gunung Lumut, Swan Slutung Village, Muara Komam District , Paser Regency , Kalimantan Selatan [- 1.41905° S, 115.966467° E, elevation ca. 300 m a.s.l.], all collected by Irvan Sidik GoogleMaps ; MZB. Amph 21487, a subadult female collected from Meratus Mountain Range, Emil Baru Village , Mantewe District , Tanah Bumbu Regency, Kalimantan Selatan on August 2010 by Dwi Susanto and Jarot Arizona ; MZB. Amph 30471–30473, three adult females collected from Production forest Aya Yayang Indonesia at Meratus Mountain Range , Mantui river , Hegar Manah village , Batang Ara district , Tabalong Regency , Kalimantan Selatan [- 1.769444° S, 115.324389° E, elevation ca. 348 m a.s.l.] by Daru Y. Setyanto between 24–27 July 2001 GoogleMaps .
Referred specimens. MZB. Amph 25435, a subadult female from Meratus Mountain Range , Amandit River , Lok Lahung Village , Loksado District , Hulu Sungai Selatan Regency , Kalimantan Selatan, Borneo ; MZB. Amph 25053, a subadult female collected from the same locality with holotype ; MZB. Amph 25262, an immature individual from Meratus Mountain Range , Desa Dua , Binuang Santang Village , Halong District , Balangan Regency , Kalimantan Selatan , Borneo on February 2009 by Irvan Sidik (- 2.400073°S, 115.742082°E, elevation 340 m a.s.l.) GoogleMaps ; MZB. Amph 30511.1−2, tadpole stage 36 and 30 from Meratus Mountain Range , Loksado village , Hulu Sungai Selatan Regency , Kalimantan Selatan, Borneo (- 2.796806° S, 115.503696° E, elevation 300 m a.s.l) GoogleMaps ; KUHE 53577 View Materials , metamorphosis stage 45 from Bario , Sarawak, Borneo .
Etymology. Specific epithet “ kalimantanensis ” is a toponym in reference to Kalimantan, Indonesian part of Borneo Island, where the type locality of the species is located.
English name. Kalimantan Horned Frog
Indonesian name. Katak-tanduk kalimantan
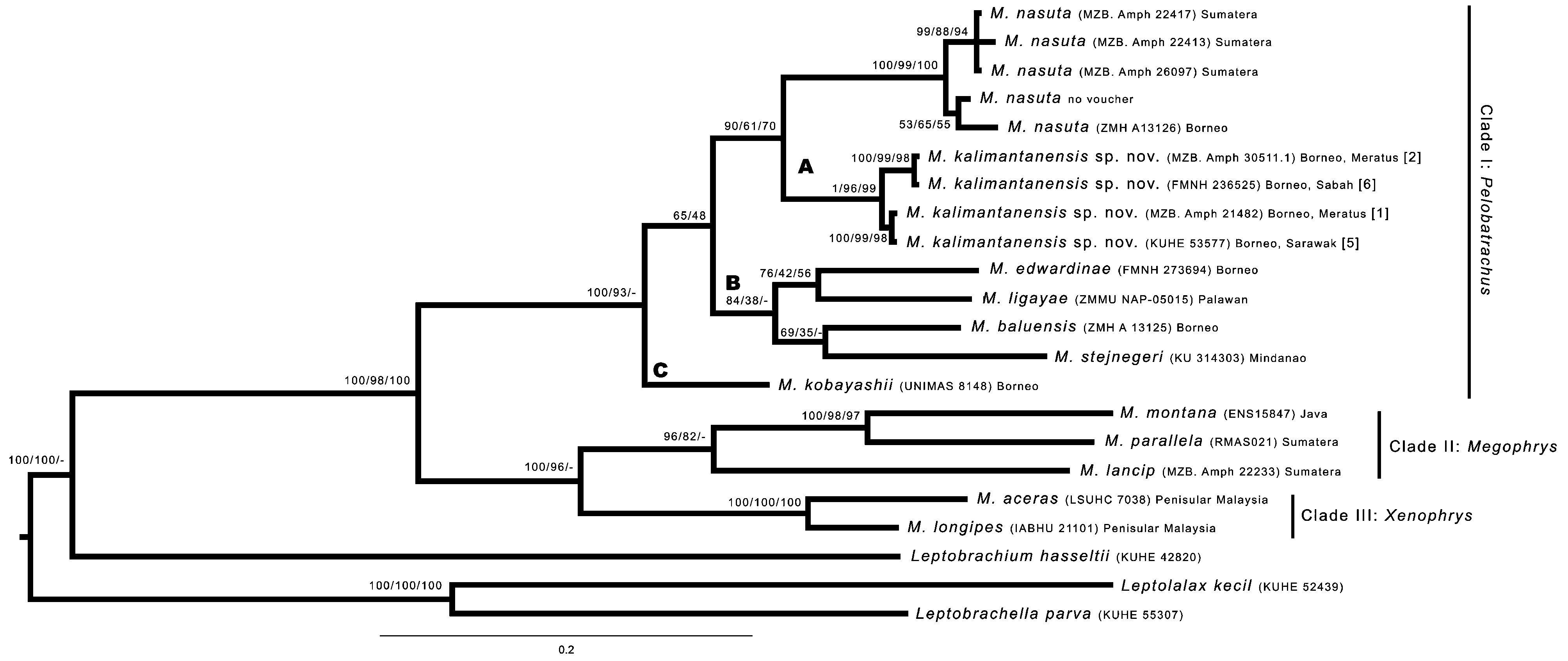
Diagnosis. We consider a new frog belonging to the genus Megophrys based on the morphology of tadpoles, having dorsally orientated umbelliform oral discs ( Smith, 1926; Lathrop, 1997; Dubois & Ohler 1998), and based on supported molecular hypothesis ( Fig. 2 View FIGURE 2 ). Megophrys kalimantanensis sp. nov. is diagnosable from its related congeners by having a large body (SVLh: 64.8–101.1 mm in adult males, 109.6–116.4 mm in adult females); acute snout with short rostral appendage (RSAL: 0.1–0.8% in males, 0.1–0.5% in females); short snout (RSLh: 11.3–12.9% HLh) protruding in lateral view; head length is shorter (RHLh: 37.2–44.1% in males, 40.3–42.5% in females) than head width (RHW: 106.9–199.9% RHLh in males, 110.2–112.7% RHLh in females); upper eyelid (UE) with short triangular, orbital appendage (EHL: 32.1–41.6% UEWh in males, 31.1–37.6% UEWh in females); tympanum distinct, vertically longer than its width; two widely separated vomerine teeth groups at posterior border of choana; absence of subarticular and outer metatarsal tubercles; a pair of dorsolateral folds extending from shoulder to groin; presence of an additional pair of lateral fold on the flank.
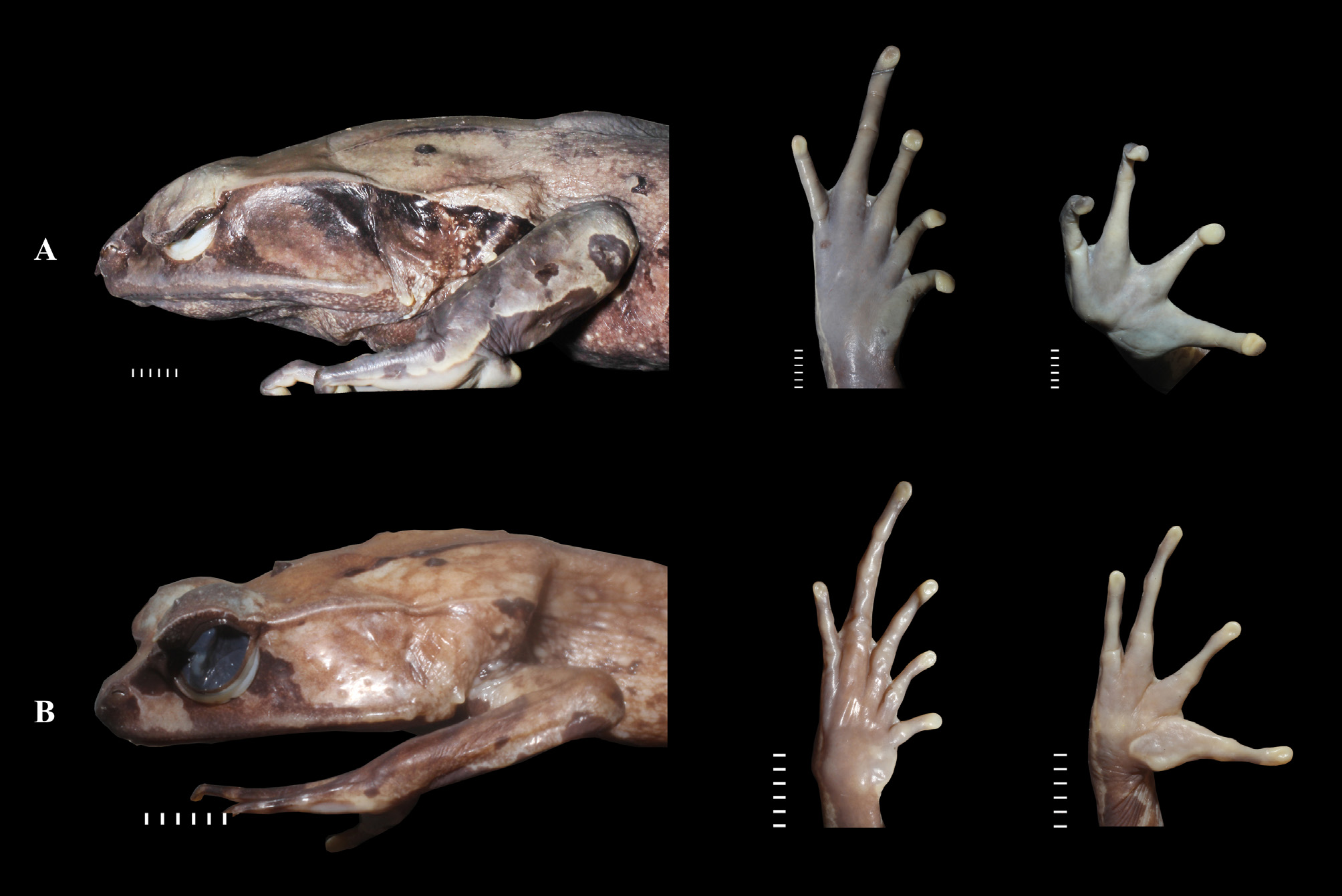
Description of Holotype (in mm). Body large (SVLh 91.0; SVL 90.8) and stocky; head depressed and broad, slightly wider (HW 38.9: 42.7% SVLh) than long (HL 33.6: 36.9% SVLh); snout protruding in lateral view, acutely pointed in dorsal view with a short dermal, rostral appendage (SAL 0.2: 0.2% SVLh), projecting beyond lower jaw; eye laterally positioned, large, more than twice and half of tympanum horizontal diameter (ED 12.0: 266.6% TDH), longer (ED 12.0: 13.2% SVLh) than snout–horn (SLh 10.4: 11.4% SVLh) and shorter than nostril to eye (NEL 5.0: 5.5% SVLh); canthus rostralis with sharp, angular ridge; lore sloping and concave; internarial distance (IND 7.1: 7.8% SVLh) about half of interorbital distance (IOD 16.8: 236.6% IND); orbital appendage about 0.37 times of total upper eyelid width (EHL 4.5: 37.2% UEWh), tip of orbital appendage acute, surface smooth scattered with small and low tubercles; tympanum distinct, oval, slightly longer vertically (TDV 4.6: 5.0% SVLh) than horizontally (TDH 4.5: 4.9% SVLh); distinct, angular supratympanic fold widened anteriorly, narrowed posteriorly, extending from behind eye, curving down around upper border of tympanum and terminating above axilla; white conical tubercles behind the supratympanic fold and front of axilla; spinous gland on corner of mouth on jaw angle; single row maxillary teeth present; vomerine teeth in two widely separated groups on level connecting posterior borders of choana; tongue mushroom-shaped, without papillae, notched posteriorly: median subgular vocal sacs with slit-like opening on each side of jaw commissures.
Forelimb slender, short (FLL 57.88: 63.5% SVLh), lower arm proximally enlarged, wider than upper arm; fingers moderately slender, with rounded and swollen tips, unwebbed and lacking lateral fringes; finger formula II<I<IV<III (fin1L 7.2: 7.8% SVLh; fin2L 6.6: 7.3% SVLh; fin4L 8.4: 9.2% SVLh; and fin3L 10.7: 11.8% SVLh); subarticular tubercles absent; outer palmar tubercle smaller (OPTL 4.9: 5.4% SVLh) than inner palmar tubercle (IPTL 6.2: 6.9% SVLh)
Hindlimb slender and moderately long (HLL 125.5: 137.9% SVLh), thigh (FML 40.5: 44.5% SVLh) longer than tibia (TL 35.1: 38.6% SVLh), tarsus (TSL 19.5: 21.4% SVLh), and foot (FL 35.9: 39.5% SVLh); toe formula being I<II<V<III<IV (Toe1L 5.8: 6.3% SVLh; Toe2L 6.1: 6.7% SVLh; Toe5L 8.4: 9.2% SVLh; Toe3L 10.3: 11.4% SVLh; and Toe4L 18.9: 20.7% SVLh); tibiotarsal articulation reaching to point anterior to tympanum; inner metatarsal tubercle moderately large (IMTL 5.3: 5.8% SVLh) but outer one absent; subarticular tubercles absent, toes with rounded and swollen tips and webbed only at base.
Dorsal skin smooth with low dense tubercles mostly on forelimb, hindlimb and lateral body; a distinct pair of dorsolateral fold, extending from shoulder on axilla region to groin. Aside from dorsolateral fold, there is an additional lateral fold on the lateral side of the body, extending from corner of supratympanic fold to groin, and about one third length of flank; vent with dermal accessory extension above cloaca; three transverse folds on lower arm; four transverse folds on hind limb, two on thigh and two on tarsus; limb skin laterally smooth, with small tubercles around groin; ventral skin smooth, wrinkled in preservative; a pair of white, conical pectoral glands, at base of axilla; a pair of white, conical femoral glands, at middle of lateral side of posterior thigh; dark brown coloured microspinules nuptial pads covering dorsal and median surfaces of distal half of metacarpal and proximal half of basal phalanx of first and second fingers and a narrow dorsomedial strip of second finger.
Colouration. In life ( Figs. 4 View FIGURE 4 , 5 View FIGURE 5 A–B), dark brown to orange-brown above, with a darker area occupying median strip of back between dorsolateral folds and sending wide branches to upper eyelid; forearm, hand, and hindlimb above darker than dorsal body with dark crossbars, three on forearm and two on thigh and tarsus, medially with a light brown streak; knee with irregular dark spot; laterally tip of snout dark brown, a light orange brown bar present behind of nares to eye, a dark bar below eye; sides of trunk unmarked; dark brown reticulum on ventrum; orange brown on throat with three longitudinal, dark stripes; palmar of hand and feet dark brown, posterior and ventral surfaces of thigh to tarsus dark brown with white spots; iris brown with dark vertical pupil. In preservative orange brown colour changed to be pale ( Fig. 5C View FIGURE 5 ).
Tadpole (stages 30 and 36 in mm). Body slender, ellipsoidal (TTL, 30 = 38.8; 36 = 47.0), about one and half as long as broad, flattened above; body (BH, 30 = 6.3: 6.2% TTL; 36 = 5.2: 11.2% TTL) slightly higher than tail muscle height (TMH, 30 = 5.8:1 15.0% TTL; 36 = 4.7: 10.0% TTL); the mouth labia dorsoterminal with a surface-oriented umbelliform oral disc; submarginal papillae are present on both sides between the oral orifice and emergination of upper and lower lip, concentrated around the margins, keratodonts are absent; head broader than trunk; eyes positioned laterally (ED, 30 = 1.7: 4.4% TTL; 36 = 1.4: 2.9% TTL); nares oval, opening laterally and dorsolaterally positioned, closer to snout than to eye (NE, 30 = 1.1: 84.6% ES; 36 = 1.6: 58.3% ES); internarial distance (IND, 30 = 3.0: 7.7% TTL; 36 = 3.6: 7.7% TTL) smaller than interorbital distance (IOD, 30 = 4.1: 10.6% TTL; 36 = 4.5: 9.6% TTL); spiracle sinistral; anus median, anal tube ending at margin and separated from ventral fin ( Fig. 8C View FIGURE 8 ); tail muscle strong about 72.5% of total length; fin beginning at base of tail; dorsal fin deeper than ventral fin, tail tapering gradually from centre to blunt tip ( Fig. 8 View FIGURE 8 A–C). The tadpole body measurement, including stage 45, is shown in table 4.
Background colour of body and tail of living specimens (stage 30 and 36) semi-transparent dark-brown with irregular white spot, slightly lighter on rostral appendage; a conspicuous pattern of dark brown and gold or orange brown pigmentation on dorsum of body and half of tail, particularly on dorsal part of head and trunk reaching laterally below eye; a shiny lateral stripe from base of oral umbeliform to pre-orbital below nares and border between rostral and rostral appendage; black eye sclera, iris dominated with gold brown with sprinkle dark spot, pupil rounded homogenously black; a pale stripe below spiracle; tail and fin laterally covered by a marbled pattern of dark brown pigmentation ( Fig. 8A View FIGURE 8 ); edges of upper and lower fins with golden iridophores, smaller on lower fin; in preserved specimens, the dark patterns fade and the skin turn transparent, and the orange gold pigmentation become pale whitish.
Orange gold pigmentation lost on specimen at stage 45 ( Fig. 8 View FIGURE 8 D–F), dorsal side of head and body dark brown, lighter on frontal to snout region, behind eye and limbs; light orange gold pattern on dorsal surface of elbow and tips of fingers and toes; limbs dorsally with dark cross bars; dorsum of tail covered with a marbled pattern of gold; ventrum of throat black with small white spot; dark brown marbled pattern on ventrum of body and limbs, While the tail ventrally marbled light orange gold; laterally, body and tail have the same color pattern as dorsum.
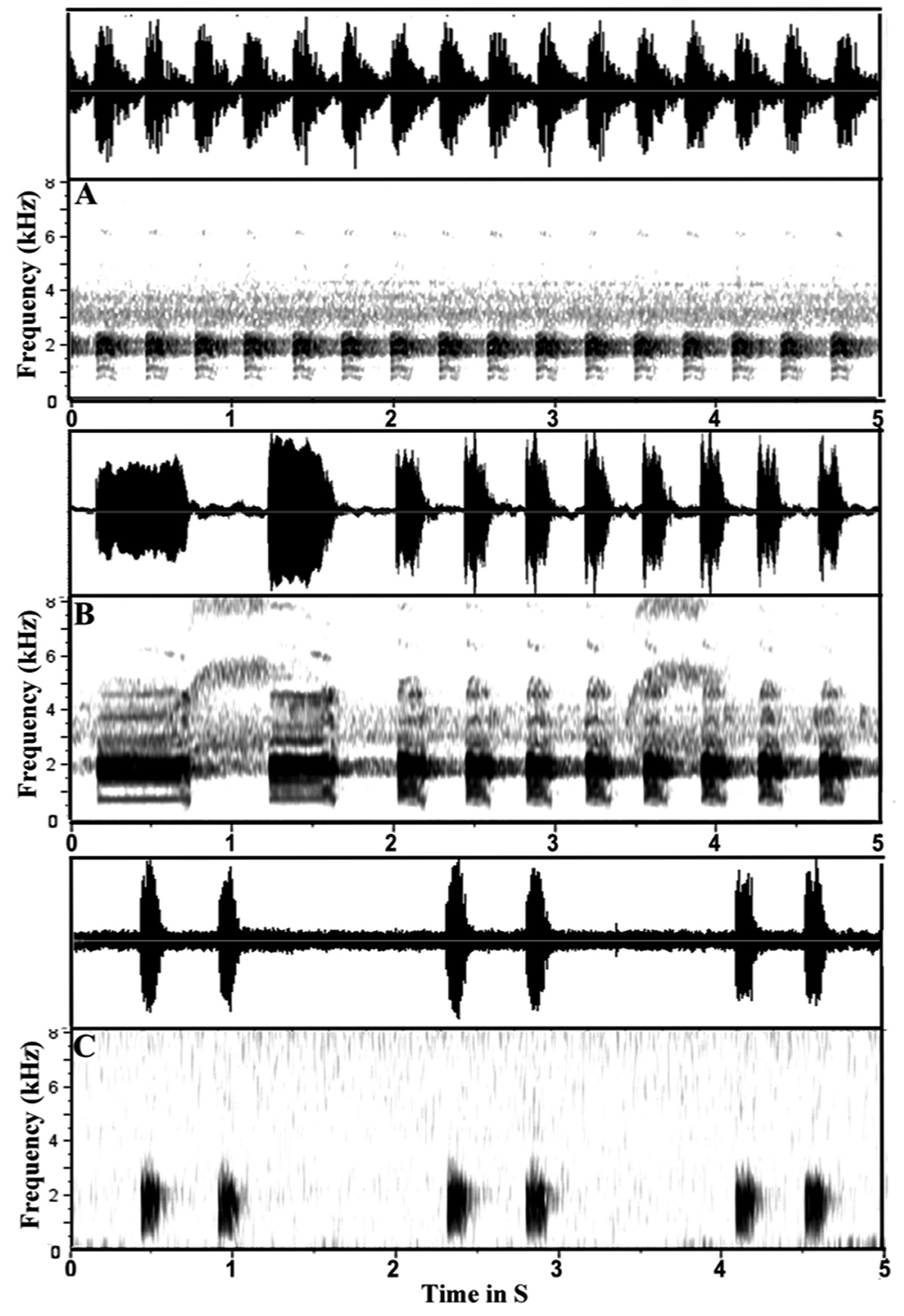
Characteristics of advertisement call ( Fig. 9 View FIGURE 9 A−B). The following description is based on the male holotype (MZB. Amph 31765). Calls contained two major types, one containing repeated short notes (type I) and another is containing repeated long notes (type II). Call type I contains 4−22 short notes with call duration of 1.1− 6.5 s (mean±SD = 3.8±6.49, n = 3), note duration 116.2−211.5 ms (mean±SD = 137.9±16.18, n = 50), internote duration 141.8−233.8 ms (mean±SD = 185.8±23.53, n = 50), note period 296.4−393.1 ms (mean±SD = 324.0±27.16, n = 50), and dominant frequency 1.8−2.3 kHz (mean±SD = 2.1±0.15, n = 50). Call type II contains 3−6 notes with call dura- tion 1.8− 5.8 s, note duration 268.7−885.6 ms (mean±SD = 536.9±195.08, n = 6), internote duration 323.7−535.1 ms (mean±SD = 434.7±75.93, n = 6), note period 679.5−1363.2 ms (mean±SD = 1009.9±26.86, n = 6) and dominant frequency 1.9−2.2 kHz (mean±SD = 2.04±0.1, n = 6). Most of calls contained only type I ( Fig. 9A View FIGURE 9 ), but one call contained both types and began with a call type II followed by the type I ( Fig. 9B View FIGURE 9 ).
Variation. Individuals of the type series are morphometrically similar. A summary of morphometric data is shown in Tables 5 View TABLE 5 & 6 View TABLE 6 together with those on the allied species, M. nasuta . In SVLh, the adult females (mean ± SD = 113.7 ± 3.19 mm, n = 4) are significantly larger than the adult males (81.4 ± 14.50 mm, n = 6). Relative lengths of rostral appendage (RSAL) slightly larger in males (median = 0.5 %) than females (0.4 %), but the variation range largely overlaps. Relative length and width of head tend smaller in males (RHLh 39.6%; RHW 44.3%) than females (RHLh 41.4%; RHW 46.1%) but overlap in range of variations in both sexes. Relative length of orbital appendage (REHL) is larger in males (median = 4.7%) than females (3.9%).
When the hindlimb is bent forward along the body, tibiotarsal joint usually reaches to the posterior corner of the eye in adult males and subadult females, but in adult females, the joint reaches the posterior of tympanum. The transverse fold on the occipital region varies in degree of development, and although most adults possess it, the holotype, two male paratypes (MZB. Amph 31766; 31767), and subadult females lack it.
Individuals are only slightly variable in coloration. Longitudinal dark brown patch on the centre of throat seen in the holotype is also found in all males and all females, but lacking in one male (MZB. Amph 25056). Dorsum in males and females is darker, but is lighter than holotype in MZB. Amph 25054; 25055; 25728. Black conical tubercles on dorsum are present in males and female paratypes but absent in holotype and two male paratypes (MZB. Amph 25054; 31766), positioned on occipital region, both side of upper tympanum and sacral regions ( Fig. 6A View FIGURE 6 ).
Secondary sex characters. Males have vocal slits on the corner of jaw, one each side and rugose dark nuptial pads covered with microspinules on the first and second fingers, positioned dorsomedially from the basis to the first subarticular joint of the finger, and enlarged lower arm.
Comparisons. Within Sundaland and Philippines Megophrys , the new species can be distinguished from the members of the subgenus Xenophrys ( M. aceras and M. longipes ) and M. dringi by the absence versus presence of Y, X or H folds directed backwards at the parietoscapular region to the axilla level ( Boulenger 1885; Boulenger, 1903; Inger et al. 1995). Molecularly the new species belongs to subgenus Pelobatrachus , however morphologically, it is difficult to differentiate it from the subgenus Megophrys , since there is no significant key character available to differentiate between these two subgenera. Based on the type of dorsolateral fold categorised by Munir et al. (2018), M. kalimantanensis sp. nov. is included in type I together with M. lancip , M. montana , M. nasuta , M. parallela , and M. stejnegeri .
Megophrys kalimantanensis sp. nov. and M. nasuta can be morphologically differentiated from all the members of the subgenus Megophrys ( M. lancip , M. montana , and M. parallela ) by presence of an additional lateral fold that are elongated from supratympanic fold to the end of the trunk ( Munir et al. 2018). This additional lateral fold is also present on M. kobayashii and M. ligayae but differs in shape. In M. kobayashii and M. ligayae the lateral fold does not extend more than two-thirds length of the trunk ( Taylor 1920; Inger 1954; Malkmus & Matsui 1997), but it continues to end of the trunk in M. kalimantanensis sp. nov. and M. nasuta . While M. kobayashii and M. ligayae have type II dorsolateral fold ( Munir et al. 2018), M. kalimantanensis sp. nov. has type I.
Megophrys kalimantanensis sp. nov. is superficially similar to the widespread species M. nasuta and has been confused with it molecularly ( Frost et al. 2006; Hamidy et al. 2012, Matsui et al. 2017a). Our molecular results support their close relationships ( Fig. 2 View FIGURE 2 ), but they can be clearly differentiated morphologically in some characteristics (see Tables 5 View TABLE 5 & 6 View TABLE 6 ). Megophrys kalimantanensis sp. nov. has a much shorter rostral appendage (RSAL 0.1–0.8 in males; 0.1–0.5 in females) than M. nasuta (RSAL 2.3–9.5 in males; 1.8–8.4 in females), shorter head (RHLh 37.2–44.1 in males and 40.3–42.5 in females vs. 40.8–53.9 in males and 42.9–57.2 in females), narrower head (RHW 41.5–48.9 in males and 45.1–47.7 in females vs 45.5–52.8 in males and 45.7–57.4 in females), shallower head (RHD 15.3–19.5 in males and 15.7–17.1 in females vs. 16.0– 25.5 in males and 15.4–24.1 in females), shorter snout (RSLh 11.3–12.9 in males and 11.2–12.5 in females vs. 9.1–21.9 in males and 12.6–21.7 in females), narrower upper eyelid (RUEWh 11.2–15.7 in males and 9.8–11.8 in females vs 14.0– 22.6 in males and 11.2–20.0 in females). Megophrys kalimantanensis sp. nov. tends to have a shorter orbital appendage (REHL 4.4–5.0 in males and 3.2–4.4 in females) than M. nasuta (7.2–12.0 in males and 4.6–12.0 in females), and narrower inner palmar tubercle than the latter (RIPTL 6.1–9.6 in males and 6.8–9.5 in females vs. 7.3–11.2 in males and 7.2–11.6 in females). The rostral and orbital appendages are acute in M. kalimantanensis sp. nov. but acuminate in M. nasuta . The orbital appendage ratio of M. kalimantanensis sp. nov. is smaller (REHL/RUEWh 32.1–41.6 in males and 31.1–37.6 in females) than M. nasuta ( 43.7–61.4 in males and 32.7–61.3 in females). These differences also apply in sub-adult specimens ( Table 5 View TABLE 5 & 6 View TABLE 6 ).
Megophrys kalimantanensis sp. nov. can be distinguished from its ally M. nasuta in the advertisement call characteristics. Calls of M. kalimantanensis sp. nov. contain two types, repeated single short and repeated single long notes versus repeated double notes in M. nasuta ( Fig. 9 View FIGURE 9 A–C), and differ from those of M. nasuta by longer call duration (1.1− 6.5 s in type I and 1.8− 5.8 s in type II vs. 0.1− 1.8 s); more notes in a call (4−22 notes in type I and 3–6 notes in type II vs. 1−3 notes); longer note duration ( type I = 116.2−211.5 ms and type II = 268.7−885.6 ms vs. 76.1−154.9 ms); shorter inter-note duration in type I (141.8−233.8 ms vs. 296.9–439.0 ms); shorter and longer note period ( type I = 296.4−393.1 ms and type II = 679.5−1363.2 ms vs. 421.8−563.8 ms); and slightly higher dominant frequency ( type I = 1.8−2.3 kHz and type II = 1.9−2.2 kHz vs. 1.7−2.2 kHz).
Distribution and Natural History. The type series was collected in Southern Borneo from the secondary to primary forests that extended from 245 to 1000 m a.s.l. in the west slope of the Meratus Mountain Range, while a paratype was collected from secondary forest of Mount Lumut at around 300 m a.s.l. There are also records of this species from northern Borneo (KUHE 53577: Bario, Sarawak; FMNH 236525: Crocker Range, Sabah). The holotype was collected from the bank about 1 m from the rocky stream ( 1 m in width). The tadpoles from Meratus Mountain were collected from a small rocky stream (about 1.5 m in width and 40 cm in depth) on the rubber plantation, which was relatively close to the human settlements.
In the beginning of rainy season ( September 2018), when the forest floor was still dry, the call of males was heard at sunset (18:00–18:30). But, this call also could be heard at midnight and even in the daytime at the end of the rainy season ( March 2019), especially during the light rain or after a heavy rain. The inguinal amplexus individuals were observed in the waterfall at 16:00 on 15 March 2019 after a heavy rain between 12:00–15:00 ( Fig. 10 View FIGURE 10 ). We found that they were mobile, sometimes submerged among the rock under the current at the shore of waterfall, about 10 m from the waterfall. Other ecological and biological information (e.g. populations, distributions, habitat requirements etc.) of the new species is unknown. The following anuran species have been found sympatrically with the new species. At the Meratus Mountain range: Ansonia leptopus Günther ; Ansonia spinulifer Mocquard ; Chalcorana raniceps Peters ; Leptobrachium abbotti Cochran ; Leptobrachium montanum Fischer ; Leptolalax gracilis Günther ; Limnonectes cf. kuhlii ; Limnonectes hikidai Matsui & Nishikawa ; Limnonectes leporinus Anderson ; Metaphrynela sundana Peters; Polypedates colletti Boulenger ; Polypedates macrotis Boulenger ; Polypedates otilophus Boulenger ; Pulchrana picturata Boulenger ; Rentapia hosii Boulenger ; Rhacophorus borneensis Matsui, Shimada , & Sudin; Rhacophorus pardalis Günther ; Staurois sp. 1; at Nyahan and Mantui rivers: Amnirana nicobariensis Stoliczka ; Ansonia leptopus ; Ansonia spinulifer ; Chalcorana megalonesa Inger, Stuart , & Iskandar; Fejervarya limnocharis Gravenhorst ; Hylarana erythraea Schlegel ; Ingerophrynus divergens Peters ; Leptobrachium abbotti ; Leptolalax gracilis ; Limnonectes cf. kuhlii ; Limnonectes cintalubang Matsui, Nishikawa , & Eto; Limnonectes finchi Inger ; Limnonectes leporinus ; Limnonectes paramacrodon Inger ; Limnonectes rhacodus Inger, Boeadi , & Taufik; Meristogenys sp. 1; Meristogenys sp. 2; Microhyla malang Matsui ; Occidozyga sumatrana Peters ; Odorrana hosii Boulenger ; Phrynoidis asper Gravenhorst ; Polypedates leucomystax Gravenhorst ; Polypedates macrotis ; Pulchrana picturata ; Rentapia hosii ; Rhacophorus gauni Inger ; Rhacophorus pardalis ; Staurois cf. tuberilinguis ; Staurois guttatus Günther.
Conservation status. Megophrys kalimantanensis sp. nov. occurs in the region of the production forest area and protected forest that faces the high pressure of illegal logging. The potential threats are habitat destruction and the harvest for commercial use as pet. This species may qualify as Vulnerable of IUCN Redlist status and needs attention for further conservation efforts.





| MZB |
Museum Zoologicum Bogoriense |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.