
Delopleurus Erichson 1847
|
publication ID |
https://doi.org/10.1080/00222933.2014.909072 |
|
DOI |
https://doi.org/10.5281/zenodo.4331143 |
|
persistent identifier |
https://treatment.plazi.org/id/F91EE008-103B-FFBE-FDF2-DAFB9AD56C0A |
|
treatment provided by |
Carolina |
|
scientific name |
Delopleurus Erichson 1847 |
| status |
|
Delopleurus Erichson 1847 View in CoL View at ENA
Type species: D. pullus Boheman 1857 View in CoL .
Delopleurus View in CoL was established by Erichson (1847) for ‘one small south African species, which differs from Coptorhina View in CoL in narrower hind legs’. Ten years later Boheman (1857) gave the name to this species, D. pullus View in CoL , and so fixed the type species of the genus. Sharp (1876) described Coptorhina parva View in CoL from northern India. Arrow (1931) moved C.parva View in CoL to Delopleurus View in CoL and described the second species from India, D.striatus View in CoL . Paulian (1934) described the third species from India, D. cardoni View in CoL . Janssens (1939) described the second species from Africa, D. gilleti , and gave a diagnostic key to all species known by then. Frey (1963) described D. janssensi View in CoL from Ethiopia. Except for this primary taxonomic literature, members of the genus were listed in the catalogues and monographs ( Péringuey 1901; Gillet 1911; Balthasar 1963; Ferreira 1972; Král and Löbl 2006; Davis et al. 2008) but few additional data were given.
Diagnosis
Delopleurus and closely related Coptorhina share somewhat rectangular metepisternum, widest in its hind part, very convex epipleural margin ( Figure 1A View Figure 1 ), and other taxonomic characters of the Sarophorus group of genera ( Frolov et al. 2008). From Coptorhina it can easily be separated by quadridentate clypeus and smaller body size ( 3.5–6.1 mm in Delopleurus and 8.0–20.0 mm in Coptorhina ).
Description
Beetles are small ( 3.5–6.1 mm), strongly convex, black or dark brown, glabrous.
Clypeus distinctly quadridentate in most species. Two medial teeth are almost always acute, lateral ones are acute to right-angled. In some species, lateral clypeal teeth are almost not separated from the lateral margin ( Figure 7A, D View Figure 7 ). Head without carinae on disc, and lacking horns or tubercles. Small carina present near inner margin of eye. Genae right-angled, indistinctly separated from clypeus. Frontoclypeal and genal suturae almost indistinct. Eyes small, their dorsal parts small, slit-shaped in some species, ventral parts larger, sub-rectangular. Distance between eye and gula approximately twice the width of eye in ventral view. Gula with longitudinal groove ( Figure 9F View Figure 9 ). Dorsal surface of clypeus rugose in most species, frons densely punctate to rugose.
Pronotum more or less trapezoidal, about two times wider than long. Anterior and lateral margins with distinct border, base with or without border. Pronotum is not excavated, without horns or ridges but with fine short longitudinal groove basally along the midline.
Elytra trapezoidal, as wide as long, shiny to opaque, with deep sinuation on lateral margins receiving convex metepisternum ( Figure 1A View Figure 1 ). Striae fine but distinct, punctate; striae 1–7 reach base of elytron, stria 8 reaches the sinuation but not the base; striae 9–10 are very close to epipleura and mostly inseparable from each other except apically. Elytral intervals flat to convex, with minute punctation. Elytra fused along suture. Scutellum not visible from above.
Wings well developed and feature a number of characters that Delopleurus share with species of the Sarophorus group of genera ( Frolov et al. 2008): brown colour except for very basal part, reduced anal area (veins J and AP 3+4 are absent), CuA widened apically along the wing margin, RA 4 does not reach the wing margin and becomes wide and indistinct apically.
Anterior tibiae have three outer teeth. Margin basad of third tooth feebly or not serrate. Spur of anterior tibia more or less bifurcated in males and apically acute to rounded and curved inwards in females. In one species ( D. naviauxi sp. nov.), anterior tibia with one small acute tooth between first outer tooth and spur ( Figure 1D View Figure 1 ). Outer margins of middle and posterior tibiae without transverse carinae, serrate. Tarsi of all legs well developed, shorter than tibiae. Claws about half the length of fifth tarsal segment.
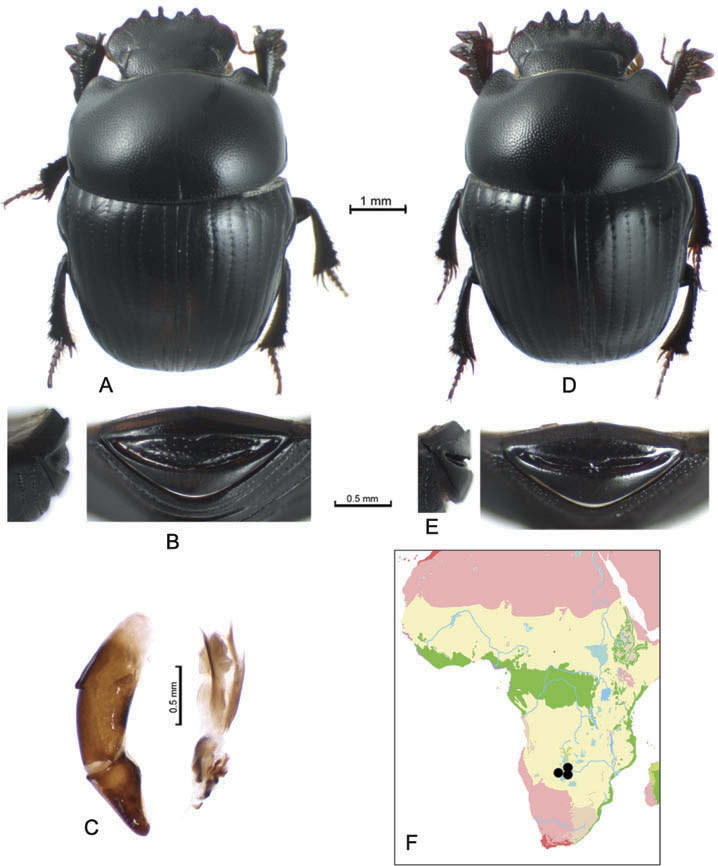
Pygidium about twice as wide as long, its sculpture is species-specific and, in some species, differs strongly from that in the other scarabaeine taxa. In some Delopleurus species basal and apical borders are relatively narrow and the disc occupies most of pygidium ( Figure 2E View Figure 2 ). In others, however, the borders are much wider and the disc is almost slit-shaped ( Figure 2B View Figure 2 ). Sculpture of pygidium is sex-dependent in some species.
Aedeagus of typical scarabaeine shape ( Figures 2F View Figure 2 , 3C View Figure 3 , 4C View Figure 4 , 5C View Figure 5 , 6C View Figure 6 , 7C View Figure 7 , 8C View Figure 8 ). Phallobase with two symmetrical tubercles. Parameres symmetrical, without setae apically, slightly distinctive in different species. Internal sac with armature consisting of a few sclerites of complex shape.
Sexual dimorphism
Males can be easily separated from females by having more or less bifurcated spurs of anterior tibiae ( Figure 1B, C View Figure 1 ). Some species differ strongly in the sculpture of the pygidium.
Diagnostic characters
Unlike other scarabaeines, Delopleurus species can be best separated by the sculpture of the pygidium. Although shape of the parameres and internal sac armature shows some species specific features, they are not considered reliable diagnostic characters. However, availability of the other characters, especially that of the pygidium, makes dissection unnecessary in routine identification.
Distribution
Delopleurus has a disjunctive range covering most of the Afrotropical Region and Indian Peninsula. The ranges of species are rather limited (except for D. gilleti Janssens ) although this may be a result of undersampling. Two regional faunas, Afrotropical and Indian, share no common species, which may indicate that they have long developed in isolation. The genus range disjunction can be best explained by the contraction of a previously very large range since the late Neogene. It is possible that in the late Miocene, when huge regions of Africa and Eurasia, including the territories of the present-day Sahara and Mediterranean, were covered by the African-type savannas, Delopleurus was widely distributed in the Eurasia. No palaeontological data exist on the past occurrence of Delopleurus in Europe and extratropical Asia, but considering very low taphonomic potential of these soil dwellers, discovery of any fossil Delopleurus seems highly improbable.
Biology
Little information is available about the biology of Delopleurus and the immature stages of the genus are unknown. In Ivory Coast, a few specimens of D. gilleti were collected on mushrooms of Termitomyces cf. schimperi along with more numerous Coptorhina nitidipennis Boheman (Yves Cambefort, pers. comm.). In Namibia ( Caprivi, Darren Mann, pers. comm.), beetles of D. darrenmanni sp. nov. were collected in mushrooms ( Termitomyces sp.) on and around termite mounds, also in association with Coptorhina ( C. nitidipennis and C.angolensis Arrow ). There were apparently Coptorhina burrows under the fungal umbrella or a few centimetres away. No smaller burrows of Delopleurus were seen however. It is possible that Delopleurus can use mushroom pieces buried by Coptorhina .
Delopleurus beetles have never been found in or under dung pads, and they have never been collected by pitfalls baited with dung or carrion [Philippe Walter collected a few specimens in traps baited with fish (Yves Cambefort, pers. comm.) but these were most probably chance captures].
Available data suggest that Delopleurus are specialist feeders of agaric mushrooms ( Agaricales ). They seem to be associated with the termitophilous genus Termitomyces , a widespread fungal genus with large fruit bodies. In the case of Delopleurus and Coptorhina , feeding on mushrooms can be a secondary adaptation from the putatively generalist saprophagy of their ancestors. Feeding on mushrooms as an adaptation to arid biotopes in beetles was hypothesized by Striganova (1980).
All Delopleurus specimens examined have well developed wings and apparently are good fliers. But no specimens have been collected at light, according to the labels and data from collectors. This, along with small eyes (typical for all members of the Sarophorus group genera), suggests that the beetles have diurnal flight activity.
Key to Delopleurus View in CoL species
Males
1. Disc of pygidium more or less densely punctate ( Figures 3B View Figure 3 , 4B View Figure 4 , 5B View Figure 5 ) or granulate with short brown setae ( Figure 2E View Figure 2 ). Species from Africa .................................. 2 Disc of pygidium almost smooth, without setae ( Figure 8B View Figure 8 ). Species from Asia ... ............................................................................................................... D. parvus View in CoL
2. Disc of pygidium rugose and with sparse brown setae ( Figure 2E View Figure 2 ) ...................... ............................................................................................. D. pubescens View in CoL sp. nov. Disc of pygidium with different sculpture, without setae. ................................. 3
3. Border of pygidium relatively narrow; punctation of pygidium disc sparser ( Figure 3B View Figure 3 ) ............................................................................................. D. pullus View in CoL
Border of pygidium thicker ( Figures 4B View Figure 4 , 6B View Figure 6 , 7B View Figure 7 ), if similar to D. pullus View in CoL then punctation of pygidium disc coarser ( Figure 5B View Figure 5 ) .............................................. 4
4. Disc of pygidium almost as long as apical border ( Figure 4B View Figure 4 ) ............................. ......................................................................................... D. darrenmanni View in CoL sp. nov. Disc of pygidium 1.5–2 times longer than apical border ................................... 5
5. Disc of pygidium feebly convex, almost in plane with basal border (in lateral view, Figure 5B View Figure 5 ) .............................................................................................. D. gilleti Disc of pygidium strongly convex, distinctly separated from basal border by a deep hollow (in lateral view, Figure 6B View Figure 6 , 7B View Figure 7 ) ..................................................... 6
6. Elytra with well-developed humeral umbones, sinuate behind umbones in dorsal view ( Figure 6A View Figure 6 ). Basal border of pygidium strongly widened medially ( Figure 6B View Figure 6 ) ........................................................................................ D. krikkeni View in CoL sp. nov. Elytra with feebly developed humeral umbones, feebly or not sinuate behind umbones in dorsal view ( Figure 7A View Figure 7 ). Basal border of pygidium slightly widened medially ( Figure 7B View Figure 7 ) .............................................................. D. fossatus View in CoL sp. nov.
Females
1. Disc of pygidium more or less densely punctate, or slit-shaped, or granulate (with short brown setae). Species from Africa ............................................................ 2 Disc of pygidium almost smooth ( Figure 8E View Figure 8 ) or granulate, without setae ( Figure 9C View Figure 9 ). Species from Asia .......................................................................... 7
2. Fore tibia with small acute tooth near apical spur ( Figure 1D View Figure 1 ). Pygidium with slitshaped pubescent disc ( Figure 2B View Figure 2 ) ....................................... D. naviauxi View in CoL sp. nov. Fore tibia without tooth near apical spur ( Figure 1C View Figure 1 ). Sculpture of pygidium different; if disc of pygidium slit-shaped then it lacks setation ( Figure 4E View Figure 4 ) .... .. 3
3. Borders of pygidium relatively narrow; disc feebly convex, granulate and pubescent ( Figure 3E View Figure 3 ) ................................................................................... D. pullus Borders View in CoL of pygidium relatively wide, disc without setation ............................... 4
4. Disc of pygidium slit-shaped, with transverse carina ( Figure 4E View Figure 4 ) ......................... ......................................................................................... D. darrenmanni View in CoL sp. nov. Disc of pygidium 1.5–2 times longer than apical border thickness ................... 5
5. Disc of pygidium feebly convex, almost in plane with basal border (in lateral view, Figure 5E View Figure 5 ) .............................................................................................. D. gilleti Disc of pygidium strongly convex, distinctly separated from basal border by a deep hollow (in lateral view, Figure 7E View Figure 7 ) ............................... D. fossatus View in CoL sp. nov.
6. Elytral striae with larger punctures ( Figure 8D View Figure 8 ). Disc of pygidium smooth to indistinctly rugose ( Figure 8E View Figure 8 ) .............................................................. D. parvus Elytral View in CoL striae with smaller punctures ( Figure 9A View Figure 9 ). Disc of pygidium granulate ( Figure 9C View Figure 9 ) .......................................................................................... D. striatus View in CoL








No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Scarabaeinae |