
Liolaemus stolzmanni, (Steindachner, 1891) (Steindachner, 1891)
|
publication ID |
https://doi.org/10.11646/zootaxa.4763.1.12 |
|
DOI |
https://doi.org/10.5281/zenodo.4323632 |
|
persistent identifier |
https://treatment.plazi.org/id/FC0887D1-7E71-5469-5FBB-FEAA9342AB02 |
|
treatment provided by |
Felipe |
|
scientific name |
Liolaemus stolzmanni |
| status |
|
courtesy of Heinz Grillitsch (Natural History Museum of Vienna) we reviewed photographs of the three types of L. stolzmanni . We examined one not type specimen and take photographs of a live individual, being to our knowledge all specimen currently placed in herpetological collections and the only and first photographic record of a live individual of this species. By courtesy of Cesar Aguilar (Museo de Historia Natural de San Marcos) we reviewed photographs of one paratype of L. insolitus Cei & Péfaur 1981 . We also examined five adult specimens of L. poconchilensis Valladares 2004 , 10 adult specimens of L. reichei and six adult specimens of L. torresi (Núñez, Navarro, Garín, Pincheira-Donoso & Meriggio 2003) . All examined specimens are listed in the Appendix I. Additional data for L. insolitus was taken from description (Cei & Péfaur 1981). Characters were taken according to Langstroth (2011) and Valladares-Faúndez et al. (2018), but supralabial scales were counted in two different ways: between the rostral scale and the mouth´s rictal and between the rostral scale to the posterior level of the subocular scale. Lorilabial scales were counted between the rostral scale (above the supralabials) to the posterior level of the subocular scale. Scales were examined under magnification lenses. Midbody scales count of L. stolzmanni types was taken from Steindachner (1891). Historical evidence was taken from the published notes of the collector of C. stolzmanni types (Stolzmann 1912, in Polish).
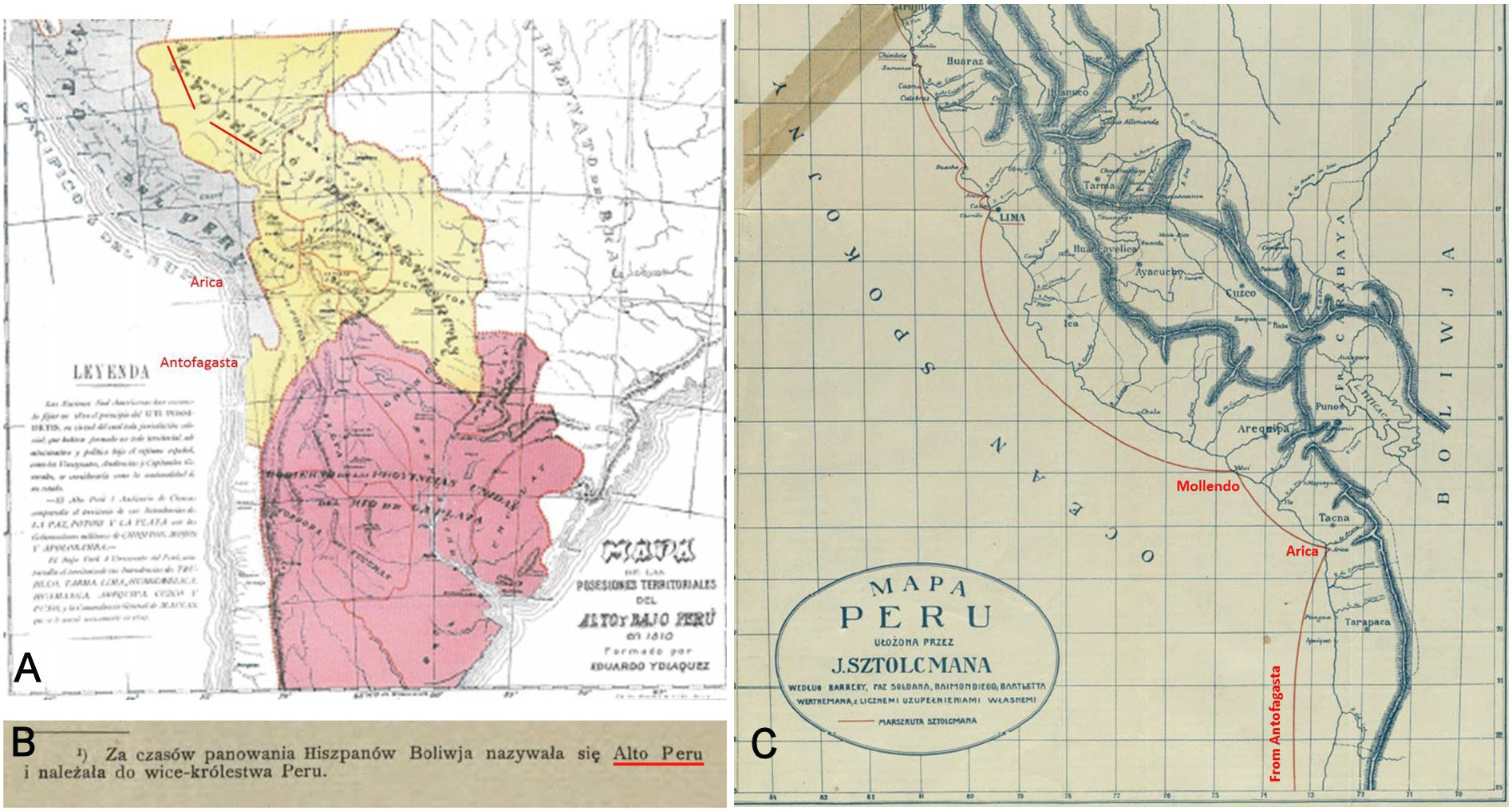
In regards to the type locality, although Langstroth (2011) brings light on several aspect of L. stolzmanni taxonomy, we partially disagree about his statements on the L. stolzmanni type locality. First, we agree in that northern Peru can be discarded because no Liolaemus occurs there ( Langstroth 2011). But we think that there is no need to suggest that Stolzmann acquired the type specimens as gifts or purchased because he carry out field campaigns to the northern Chile (southern Bolivia and Peru at 1875) and in the current southern Peru, in places in which species of “prhynosaurid” lizards have been recorded and then it can be concluded that specimens were collect by himself as was pointed out by Steindachner (1891) .
Jan Stolzmann went to South America two times, one ( 1875–1881) traveled through Chile, Bolivia and Peru and another ( 1882–1884) traveled through Ecuador ( Mierzwa-Szymkowiak & Breure 2017). This second travel can be discarded because no Liolaemus occurs in Ecuador ( Abdala & Quinteros 2014). Stolzmann (1912) described in detail his first South American travel. He declares that arrived to the Valparaíso port ( Chile) from Warsaw in 1875 and after that he traveled by sea to the Antofagasta port (currently Chile but formerly Bolivia) touching land in October 26 (p. 27). It is not stated how many days he was in Antofagasta , but he provides a detailed description of geography and industrial activities (smelting). Remarkably , Stolzmann (1912) clarify as footnote in the page 28, that Antofagasta port is placed in Bolivia (currently in Chile), and stated (in Spanish ) that it was formerly known as “Alto Peru” (translated as High Peru in English or Hoch Peru in German ), the type locality of L. stolzmanni according to Steindachner (1891). Later , he traveled by sea to Arica port (currently Chile but formerly Peru) performing a basic description of the environment. However , Stolzmann (1912, p. 29) stated that he was in Arica only few hours, and then the collection of Liolaemus seems to be unlikely. Finally , he traveled by sea to Mollendo port in Peru, noted as Islay in the map by Stolzmann (1912), and go to Lima port ( Peru), where he began his journey by northern Peru ( Fig. 1 View FIGURE 1 ) .
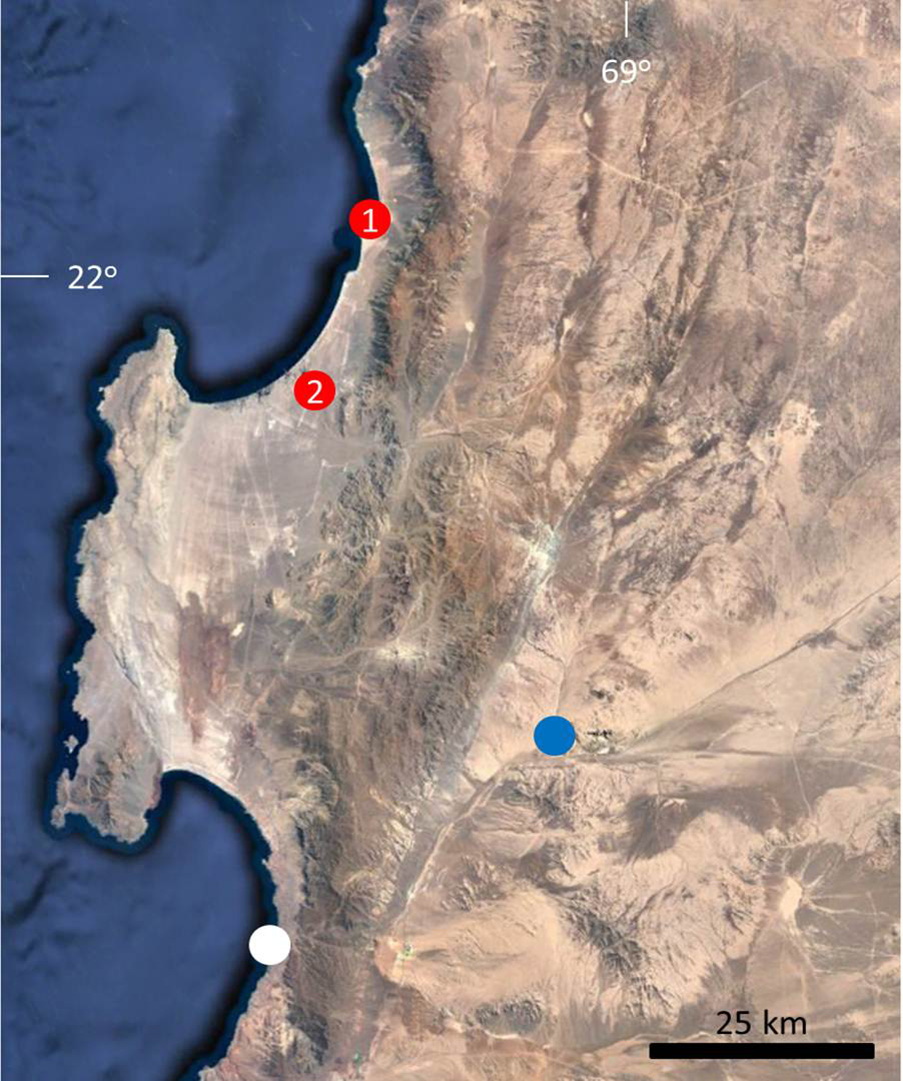
At this point, it is clear that the type locality of L. stolzmanni is in the surroundings of Antofagasta, because the reference to “Alto Peru” (High Peru) made by the collector (Stolzmann 1912) and the mention of Hoch Peru (High Peru) as the type locality by the species descriptor ( Steindachner 1891). In the aim of confirm this, we carried out a field campaign to the surroundings of Antofagasta in Marsh 7–8, 2018, and find one specimen assignable to L. stolzmanni in the desert near Mejillones ( 23º05`S – 70º21`W) at 60 km NE from Antofagasta ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 ) GoogleMaps and additionally we reviewed a another specimen of L. stolzmanni from Hornitos, MNHNCL 4620 ( 22º56`S – 70º18`W, Núñez et al. 1998; Núñez & Gálvez 2015) at 75 km NE from Antofagasta GoogleMaps . We propose to restrict the type locality of L. stolzmanni to transect between Antofagasta and Mejillones, Chile, because Mejillones is the record most geographically closer to Antofagasta . We remark that at date, this species has been not recorded in Antofagasta itself, then, probably Stolzmann collected L. stolzmanni in an excursion carry out from Antofagasta .
Moreover, we discard that Stolzmann`s collection of L. stolzmanni specimens was made in other place of his field trip, because although there are three places of his journey in which “prhynosaurid” lizards can be found, all them can be discarded: 1. Although L. torresi occurs in Mantos Blancos at 35 NE km from Antofagasta ( Riveros-Riffo & Torres-Mura 2015), it can be easily differentiated from L. stolzmanni , because L. torresi has more scales around midbody (60–68) than L. stolzmanni (approximately 50 on the types, Steindachner 1891; 45 on MNHNCL 4620); and L. torresi has the tail larger than the body, whereas the tail is shorter than the body on L. stolzmanni . 2. Although L. poconchilensis , occurs in Poconchile, 25 E km from Arica ( Valladares 2004), both can be easily differentiated because L. poconchilensis has more scales around midbody (56–62) than L. stolzmanni , and has bluish spots dispersed on the dorsum, whereas this trait is absent in L. stolzmanni . 3. Although L. insolitus occurs in the surroundings of Mollendo (Cei & Péfaur 1981), females of L. insolitus have precloacal pores, whereas the female paralectotype of L. stolzmanni has no precloacal pores. Moreover, as was noted by Langstroth (2011), both species have different color pattern, because L. insolitus has series of large dorsal dark patches with lighter shades at the center and bluish spots dispersed on the dorsum, whereas L. stolzmanni has series of small dorsal dark spots without lighter shades at the center and without bluish spots dispersed on the dorsum.
In regards to L. reichei we found that both species are very similar and can be differentiated mainly by the snout vent length (SVL). Whereas L. stolzmanni has a maximum SVL = 57.0 mm (n = 4, all known specimens), L. reichei has a maximum SVL = 50.8 mm (n = 3, Valladares-Faúndez et al. 2018; n = 10, our data). Two hypotheses arise: 1. Both species are cryptic and can be diagnosed based solely in their sizes or, 2. Both species are conspecific and their different sizes can be explained by a geographic variation, then, L. stolzmanni is a senior synonym. This should be tested through DNA phylogenetic analysis in a further study, but for the moment, and given that both species are allopatric with more than 360 km of distance between type localities, we recommend keep both as valid species to maintain the taxonomic stability as was proposed by Valladares-Faúndez et al. (2018).
On the other hand, we did not found differences in the dorsal color pattern between L. stolzmanni and L. reichei . Both have light brown dorsal color with 5–7 series of small dorsal black spots without lighter shades at the center, running from the shoulders to the base of the tail, with yellowish or orange inconspicuous transversal stripes between these, and absence of bluish spots dispersed on the dorsum ( Fig. 4 View FIGURE 4 ). We disagree with Valladares-Faúndez et al. (2018) in regards to some of the characters that they proposed as diagnostic between these species: 1. According to Valladares-Faúndez et al. (2018) both species differ on the number of supralabials scales (9 on L. reichei vs 6 on L. stolzmanni ). Apparently, this difference is due to the use of different methodologies for the scale counts. The neotype of L. reichei has nine supralabials between the rostral and the mouth´s rictal whereas has six between the rostral and the posterior level of the eye ( Valladares-Faúndez et al. 2018, Figure 3B View FIGURE 3 ). Besides, there are 9–10 supralabials between the rostral and the mouth rictal on L. stolzmanni and six between the rostral and the posterior level of the eye; in similar way, there are 8–10 and six in our sample of L. reichei ; then this feature cannot be considered as diagnostic ( Fig. 5 View FIGURE 5 ) because both species overlap these scale counts if the supralabials are counted in the same way. 2. Specimens L. reichei reviewed by Valladares-Faúndez et al. (2018) have the interparietal and both parietal scales large and well formed, as in our sample of L. reichei . However, Valladares-Faúndez et al. (2018) point out that these scales are relatively small, fragmented and irregular in L. stolzmanni , but it is not observed in none of the three type specimens of L. stolzmanni , and then it is not a diagnostic character ( Fig. 5 View FIGURE 5 ). 3. According to Valladares-Faúndez et al. (2018) both species differ in the number of lorilabials scales (eight in L. reichei vs five in L. stolzmanni ). We found 7–8 lorilabials counted between the rostral and the level of the posterior border of the subocular scale in L. stolzmanni (not counted in NMW 13580:3) and the same variation was found in our sample of L. reichei , so this character cannot be stated as diagnostic ( Fig. 5 View FIGURE 5 ). 4. According to Valladares-Faúndez et al. (2018) the ventral side is strongly variegated on L. reichei while is white, immaculate or lightly variegated in L. stolzmanni . This character cannot be considered as diagnostic because we found specimens of L. reichei with whitish immaculate ventral color. 5. According to Valladares-Faúndez et al. (2018) the lateral nuchal skin folds is well-developed and complex in L. reichei whereas this feature is not well developed on L. stolzmanni . However, our sample of L. reichei do not shows a difference for this feature in regards to L. stolzmanni .



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.