
Somaovalis sinestigma Opitz, 2016
|
publication ID |
https://doi.org/10.1649/072.070.0123 |
|
publication LSID |
lsid:zoobank.org:pub:1DAECF4D-BF16-4E97-AC0C-D2EF2CFEED57 |
|
persistent identifier |
https://treatment.plazi.org/id/FD71A26D-FFB6-0E21-FD03-4BA573414A5D |
|
treatment provided by |
Diego |
|
scientific name |
Somaovalis sinestigma Opitz |
| status |
sp. nov. |
Somaovalis sinestigma Opitz , new species ( Figs. 6, 13 View Figs , 32 View Figs , 39 View Fig , 40 View Fig )
Holotype. ♀. BRAZIL: Rondonia, 62 km SW Ariquemes, Fzda. Rancho Grande, X-8–1993. A second label reads: CW & LB O’ Brien , at merc. vap. & U V light. ( FSCA).
Paratypes. Two specimens. BRAZIL: Rondonia, 62 km SW Ariquemes, Fzda. Rancho Grande , X-10- 1993, C. W. & L. B. O’ Brien ( WOPC, 1). BRAZIL :
Rondonia, 62 km SW Ariquemes, Fzda. Rancho Grande , X-13-1993, C. W & L. B. O’ Brien ( WOPC, 1) .
Diagnosis. The black color of the elytral disc will conveniently distinguish the members of this species from congeneric specimens of S. sexnaeva , whose elytra have six pale markings.
Description. Size: Length 4.5 mm; width 1.5 mm. Form: As in Fig. 32 View Figs . Color: Cranium, pronotum, pterothorax, elytra, and abdomen black, mouthparts, antennae, and legs testaceous. Head: Cranium coarsely punctate, frons wider than width of eye (FW/EW = 30/15); funicular antennomeres filiform, antennomeres 9 and 10 subtriangular, antennomere 11 subovoid ( Fig. 6 View Figs ). Thorax: Pronotum oblong ( Fig. 13 View Figs ) (PL/PW = 68/55), coarsely punctate, without lateral tubercles; elytral disc sculptured with 8 asetiferously punctate striae that end at elytral apical 2/3 (EL/EW = 180/50), interstitial spaces large and smooth. Abdomen: Female pygidium not incised at middle of anterior margin;
Variation. The beetles before me are quite homogeneous.
Natural History. The available specimens were collected in October, one with a combination trap involving mercury vapor/ultraviolet light.
Distribution. This species is known only from Rondonia in Brazil ( Fig. 39 View Fig ).
Etymology. The specific epithet sinestigma is a compound name that stems from the Latin sine (= without) and the Greek stigma (= mark). I refer to the coloration of the elytral disc.
EVOLUTIONARY CONSIDERATIONS
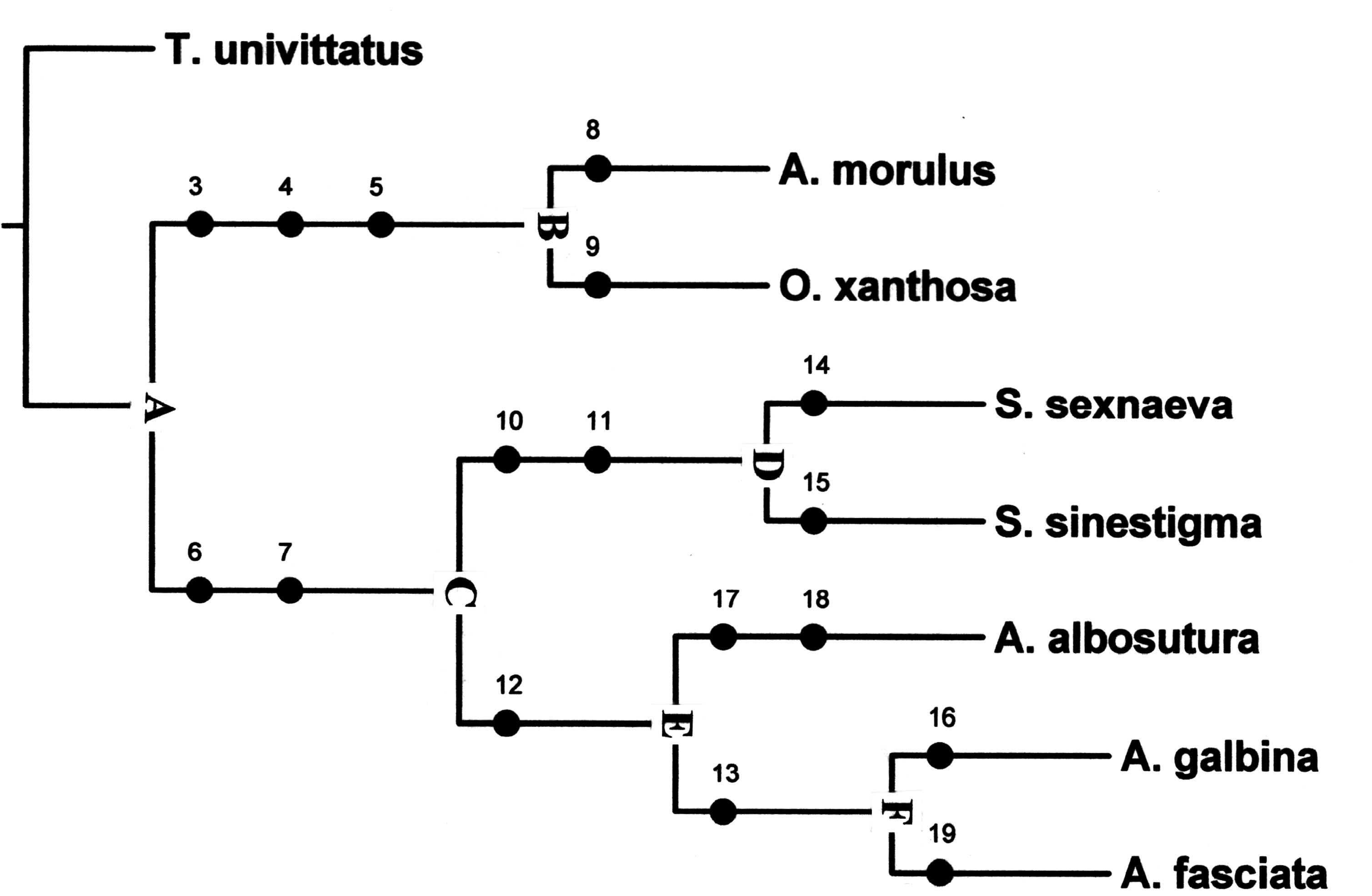
A detailed analysis of species-level morphology enabled a phylogenetic hypothesis that includes all of the species under consideration. Often, I have alluded to the scarcity of species-level characteristics for hypothesizing evolutionary history in the Cleridae ( Opitz 2009) . However, during this study I was able to find structural and color type of apotypies worthy of phylogenetic considerations at the species level. The character states listed in Table 1 were analyzed via Winclada ( Nixon 2002) in combination with NONA ( Goloboff 2003), which yielded one tree ( Fig. 40 View Fig ) involving 20 steps, index of consistency of 100, and an index of retention of 100. This species-level hypothesis is, of course, considered tentative until more data is discovered or following the discovery of new species relevant to the genera included herein.
Remarks about the biogeographical and phylogenetic character states of the species involved in this work must be considered tentative in view of the few distributional records available ( Fig. 39 View Fig ). This paucity of material may be a reflection of collecting bias and/or a general unawareness of the presence of specimens in world collections. Whatever the case, it seems likely that Pleistocene climatic changes ( Vuilleumier 1971) had a dramatic effect on the distribution of these seemingly highland species ( Table 2). That the genera in question, and their species, are of probably recent origin is suggested by their absence in Middle America, and it is likely that they evolved after the Andean orogeny during the Miocene ( Ford 2006), which established a formidable barrier to northern dispersal.
The known distributional evidence suggests that the ancestor of Ardearsus , Abiliella , Ochracea , and Somaovalis evolved in South America, perhaps in the region designated as the Altiplano Complex ( Opitz 2005 ). Three of the four genera are found in these South American Highlands. My hypothesis of the phylogenetic relationships among the four genera is depicted in Fig. 40. I View Fig hypothesize that the ancestor of these genera would have been characterized as follows: funicular antennomeres subfiliform, capitulum lax, pronotum oblong, elytral disc with 10 striae of asetiferous punctures, epipleural fold obliquely positioned, and tibial spur formula 0-1-1. This progenitor diverged towards ancestor B, in which the pronotum became transverse and the elytral asetiferous punctures were lost. In the complementary stock, which led to ancestor C, the clypeus became very setose and two striae of asetiferous punctures were lost. Progenitor B diversified to evolve A. morulus and O. xanthosa . The bifurcation of ancestor C produced ancestor D, in which the body form became subovoid and the elytral asetiferous punctures ended at elytral apical fourth. Progenitor D produced S. sexnaevea and S. sinestigma . In the sister taxon of D, ancestor E, the asetiferous punctures became binodal. Ancestor E proliferated A. albosutura , and the complementary stock that led to ancestor F, which is characterized by having the spicular apodemes fused at the posterior limits. Ancestor F then diversified to produce A. galbina and the more widely distributed A. fasciata .



| FSCA |
Florida State Collection of Arthropods, The Museum of Entomology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |