
Alonopsis elongata ( Sars, 1861 )
|
publication ID |
https://doi.org/ 10.5281/zenodo.203597 |
|
DOI |
https://doi.org/10.5281/zenodo.5671945 |
|
persistent identifier |
https://treatment.plazi.org/id/3A2BA70A-FF89-F841-FF45-FAFA180078AD |
|
treatment provided by |
Plazi |
|
scientific name |
Alonopsis elongata ( Sars, 1861 ) |
| status |
|
Alonopsis elongata ( Sars, 1861)
Leydig, 1860: 219–221, Pl. 9: figs 66–67 ( Lynceus macrourus ); Sars, 1861: 161 ( Alona ); Schoedler, 1863: 33 ( Acroperus intermedius ); Müller, 1867: 170–171, Pl. 4: fig. 28 ( elongata ); Lilljeborg, 1901: 434–440, Pl. 65: figs 5–20; Smirnov, 1966: 114–134, figs 1–10 ( Acroperus ); 1971: 413–415, figs 500–510 ( Acroperus ); Flössner, 1972: 335–339, fig. 124 ( Acroperus ); Negrea, 1983: 304–306, fig. 124; Flössner, 2000: 335–339, fig. 124.
Type locality. Lake Sognsvand (now Songsvann), the vicinity of Oslo, Norway.
Possible type material. Tube GOS F12424; slide GOS F9045 (locality is not specified, both labelled just as “ Norway ”, which is typical for material studied by Sars at this time), Zoological Museum of Oslo University, Norway.
Material studied: several specimens from the type locality, 0 9.2003, coll. A.Y. Sinev; over 100 parthenogenetic females, numerous ephippial females and males from Russia, Murmansk Area, Khibiny Mountains, Lake Malyi Vud'yavr, 11.08.2000, coll. A.Y. Sinev; over 50 parthenogenetic females from Russia, Karelia Republic, vicinity of Moscow State University White Sea Biological Station, lake Krugloe, 16.07.2008, coll. A.Y. Sinev; over 50 parthenogenetic females from Russia, Yaroslavl Area, Uglich Reservoir, 23.07.1962, coll. N. N. Smirnov, personal collection of A.A. Kotov (Severtsov Institute of Ecology and Evolution, Moscow, Russia), AAK-1999-090; 12 females from Russia, Tomsk Area, Lake Bol'shoe Purul'do, near locality Kharsk, coll. A. A. Kotov, personal collection of A.A. Kotov (Severtsov Institute of Ecology and Evolution, Moscow, Russia), AAK 2005-279.
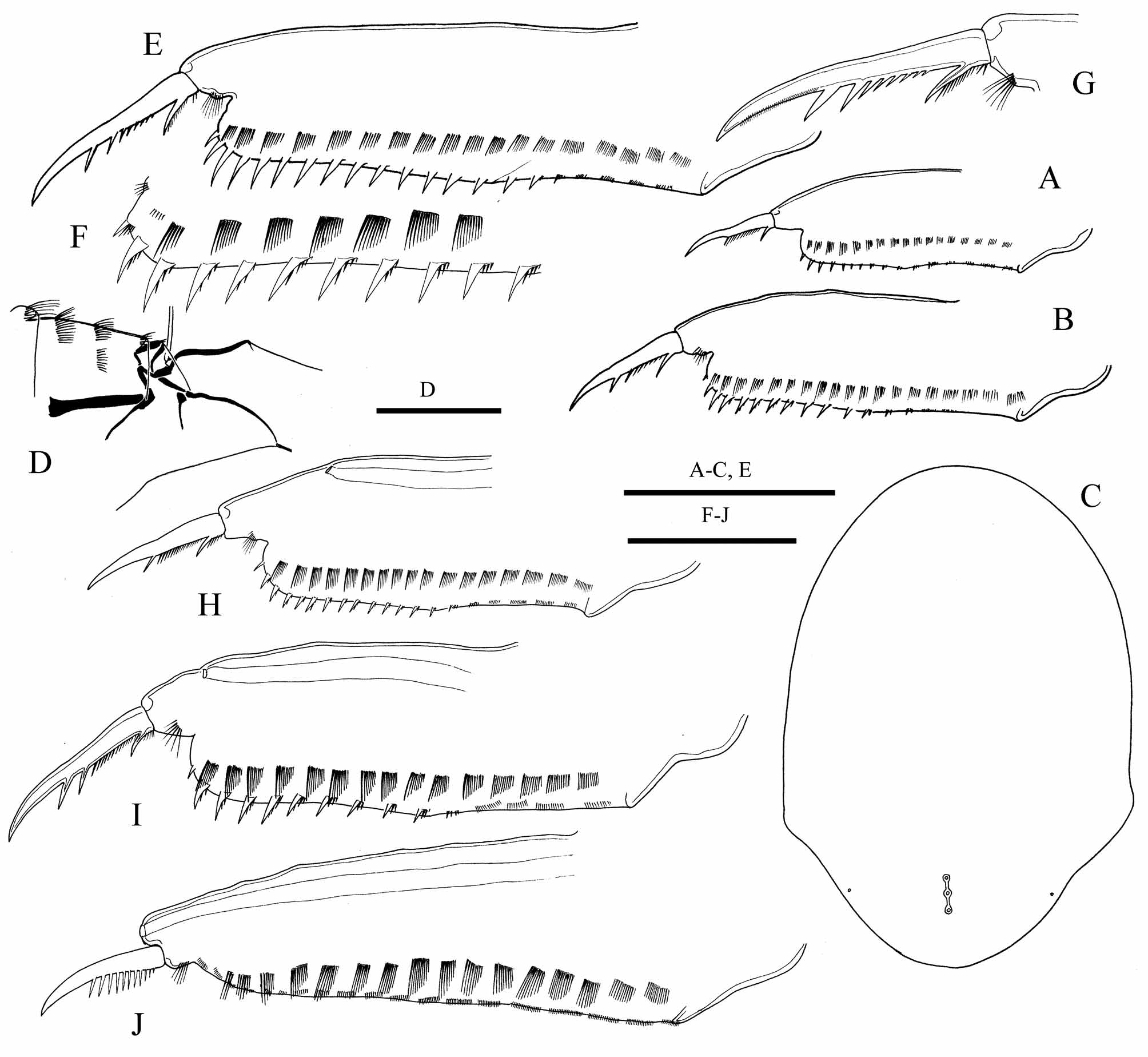
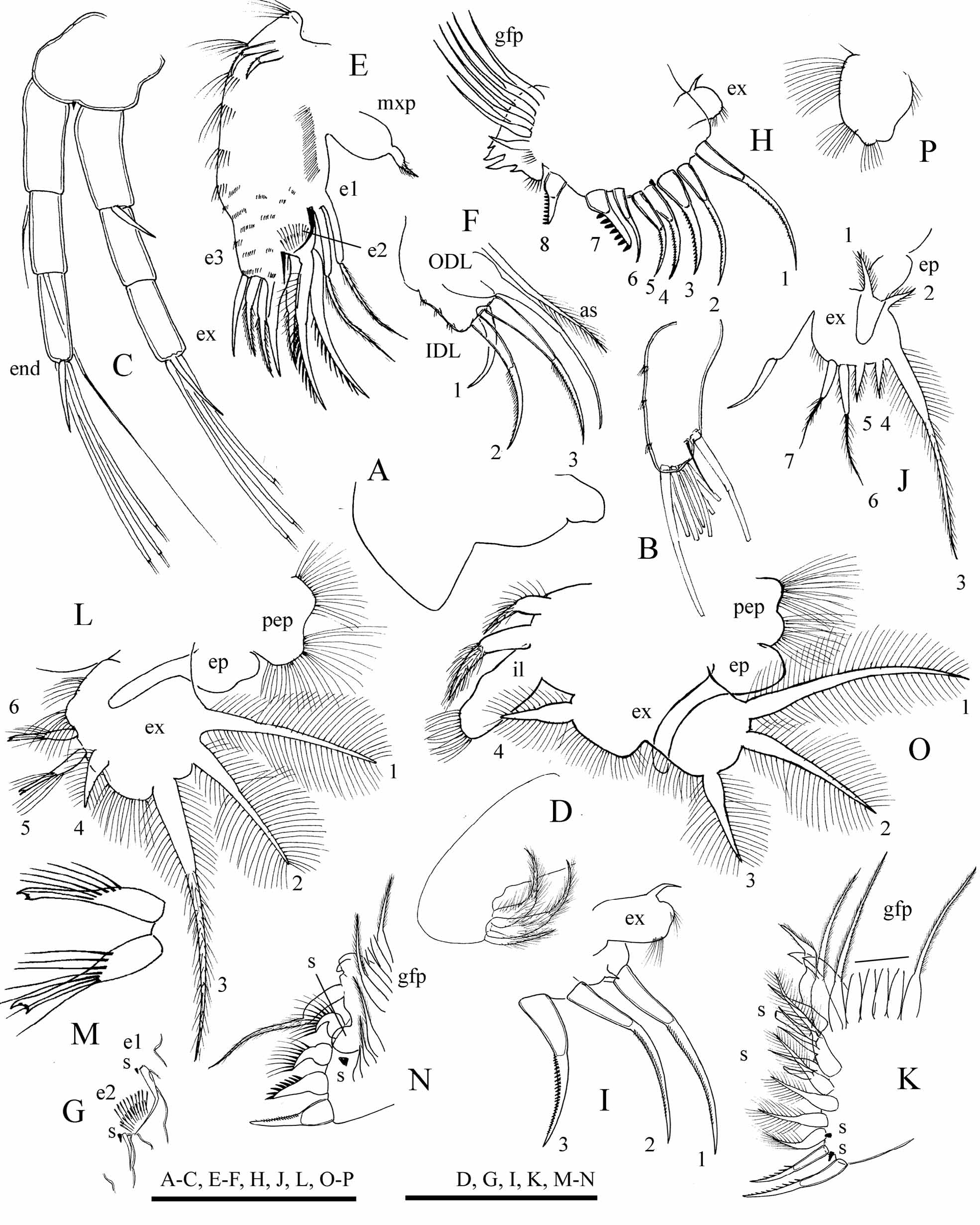
Description. Parthenogenetic female. Body ( Fig. 1 View FIGURE 1 A–D, 2A, C) height-length ratio 0.61–0.65. Valves ( Fig. 1 View FIGURE 1 E, 2D), as for genus. Head typical of the genus, head shield ( Fig. 3 View FIGURE 3 C) with broadly rounded anterior margin, with no defined rostrum. Head pores as for genus ( Fig. 2 View FIGURE 2 B, 3C), IP/PP ratio about 1.3–2.0 in adult. Labrum ( Fig. 4 View FIGURE 4 A), thorax, abdomen and postabdomen ( Fig. 3 View FIGURE 3 A–B, E–G) as for genus.
Antennule ( Fig. 4 View FIGURE 4 B), antenna ( Fig. 4 View FIGURE 4 C) and limb I ( Fig. 4 View FIGURE 4 E–G) as for genus. Limb II ( Fig. 4 View FIGURE 4 H–I) as for genus, with scraper 7 significantly larger than scraper 8, armed with 7–9 very robust, massive denticles. Limbs III ( Fig. 4 View FIGURE 4 J–K), VI ( Fig. 4 View FIGURE 4 L–N), and V ( Fig. 4 View FIGURE 4 O) as for genus; incursion between lobes of exopodite V as a right angle. Limb VI ( Fig. 4 View FIGURE 4 P) as for genus.
Ephippial female ( Fig. 1 View FIGURE 1 F) with body slightly higher than in parthenogenetic female, ephippium dark yellowbrown, without prominent sculpture.
Male. General shape of juvenile males of instar I ( Fig. 1 View FIGURE 1 G) and II ( Fig. 1 View FIGURE 1 H) similar to that of juvenile females of same instar; instar II males smaller than females of same instar. General shape of adult males ( Fig. 1 View FIGURE 1 I, 2J) similar to that of instar II juvenile females, body height/body length = 0.63–0.65. Ocellus and eye of same size as in female.
Postabdomen. In juvenile instar I males, similar to that in juvenile females ( Fig. 3 View FIGURE 3 H), with sperm duct openings located before the middle of ventral margin. In instar II juvenile males ( Fig 3 View FIGURE 3 I), shorter than that in female. Gonopores located close to the end of postabdomen. Armament of postabdomen and postabdominal claw same as in female in both juvenile instars. In adult male, postabdomen ( Fig. 2 View FIGURE 2 K, 3J) narrower than in female, narrowing distally and not curved. Postanal angle not defined, preanal angle obtuse. Distal part of postabdomen 4 times longer than preanal. Sperm duct openings at the end of postabdomen above the base of postabdominal claws. Clusters of short setules in place of marginal denticles, lateral fascicles of setules same as in female. Postabdominal claw two times shorter than that of female, curved, without basal spine, with pecten of about 10 long spines, exceeding the distalmost the width of claw base.
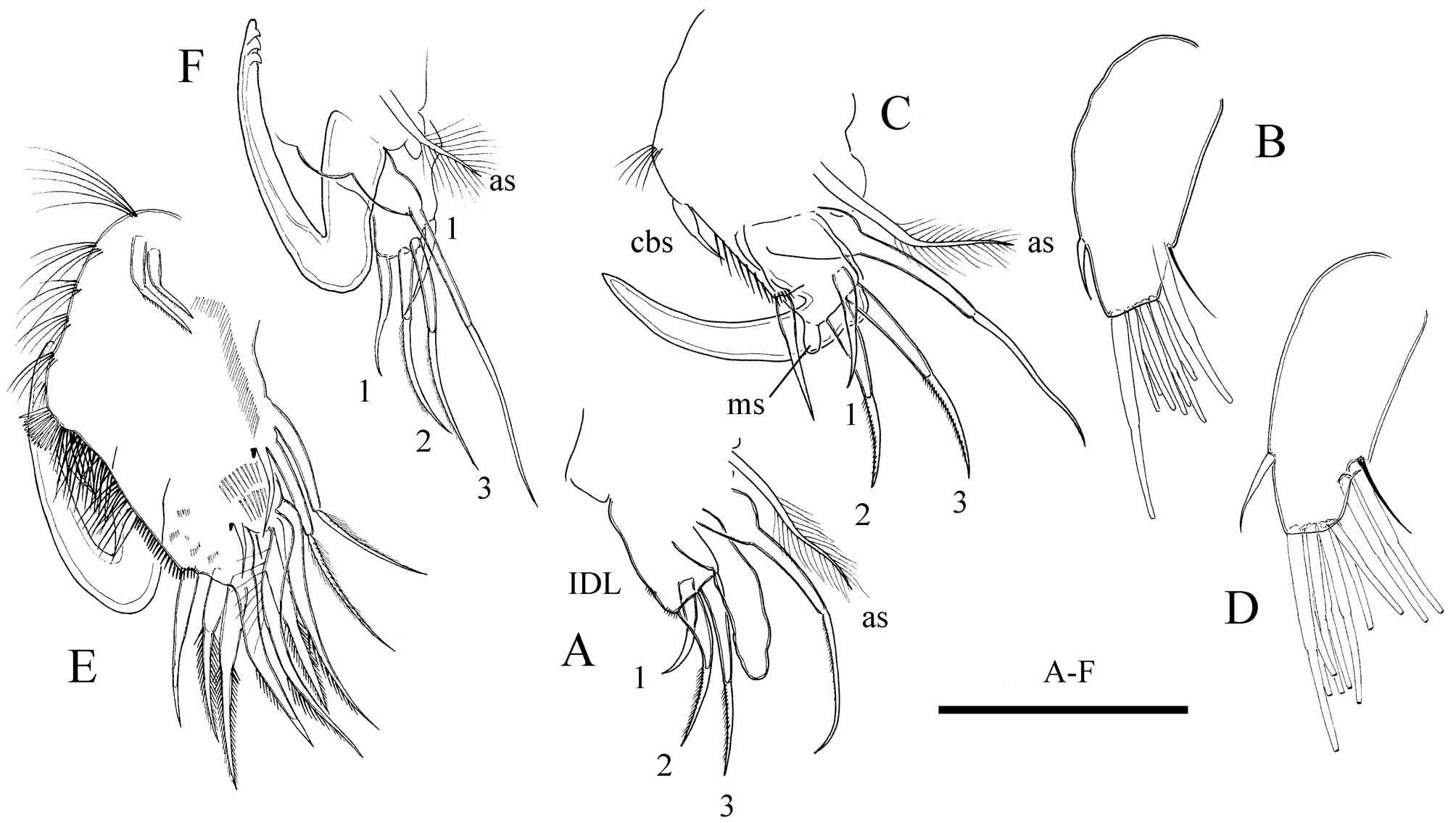
Antennule. In instar I male, same as in female. In instar II male antennule ( Fig. 5 View FIGURE 5 B) broader than in female, with an anlage of male seta, aesthetascs same as in female. In adult male antennule shorter than in female ( Fig. 5 View FIGURE 5 D), with 10 terminal and 2 lateral aesthetascs. Male seta arising at 1/4 length from tip, reaching to the end of antennule.
Thoracic limb I. In instar I male with a short anlage of copulatory hook, IDL same as in female ( Fig. 5 View FIGURE 5 A). In instar II male, copulatory hook curved ( Fig. 5 View FIGURE 5 C). Ventral face of limb with anlage of copulatory brush seta and a row of about 8 short setules below it. IDL with anlage of male seta, and other setae same as in female. In adult male, limb I stouter than that of female ( Fig. 5 View FIGURE 5 E–F), with V-shaped copulatory hook. Copulatory brush present; about 30 irregularly spaced long setules on ventral face of limb below them, following by 20 much shorter setules in regular row. IDL seta 1 present, setae 2 and 3 subequal in length, much thinner than in female; male seta thick, about half as long as seta 3.
Size. In studied material, length of females of juvenile instar I— 0.43–0.47 mm; juvenile instar II— 0.49–0.57 mm; adult female— 0.59–0.91 mm (according to literature—up to 1.1 mm). Length of male of juvenile instar I— 0.44–0.46 mm; instar II— 0.47–0.51 mm; adult male— 0.54–0.57 mm (according to literature, up to 0.6 mm).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Genus |