
Ameerega imasmari, Brown & Siu-Ting & May & Twomey & Guillory & Deutsch & Chávez, 2019
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4712.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:FC7A2CB9-5541-42AA-A1CA-6EF6BFBA0940 |
|
persistent identifier |
https://treatment.plazi.org/id/682D995B-E574-8D27-FF3C-FB3E5257FE53 |
|
treatment provided by |
Plazi |
|
scientific name |
Ameerega imasmari |
| status |
sp. nov. |
Ameerega imasmari sp. nov.
Riddle poison frog
Phyllobates pictus (non Bibron): Silverstone 1976 p. 40–41, pattern 5 (partim), trail between Satipo and Puerto Ocopa (Museum of Comparative Zoology, Havard, 24433).
Dendrobates pictus (non Bibron): Myers, Daly & Malkin 1978, p. 332 (by implication).
Epipedobates hahneli: Roberts et al. 2006 View in CoL . Sample “P1” (by implication).
Ameerega View in CoL sp. “Ivochote”: Siu Ting 2012, p. 17–22.; Guillory et al. 2020 p. 3–10.
Holotype. CORBIDI 20575 View Materials (field number JLB-WS-033), an adult male collected by Jason L. Brown, Wilson X. Guillory, and Brian Widmer in the region of Pasco, 1.5 km E of Satipo town in small patch of secondary forest adjacent to Río Satipo , Peru, 605 m elevation, 11° 14′ 14.892 S, 74° 37′ 3.576 W, 20 June 2018 ( Figs. 1 View FIGURE 1 & 4 View FIGURE 4 ). GoogleMaps
Paratypes. UMMZ 2449821–244984 View Materials and MUSM-H 35670–35673 collected by Joanna Larson and Consuelo Alarcón, in the Cusco region, Peru, Villa Carmen Biological Station and Reserve, Pillcopata, Peru, 519 m elevation, 12° 53′ 45.01 S, 71° 24′ 21.69 W, collected between 16 January 2016 and 20 February 2016 (Supp. Figs 2 View FIGURE 2 E–F, 4A–E) GoogleMaps
Etymology. The species name is formed as an adjective, derived from the Quechua word ‘imasmari’ which means ‘riddle’. The epithet is in reference to cryptic color pattern of this species, which is similar to that of Ameerega hahneli sensu stricto and A. picta sensu stricto, two species that co-occur with A. imasmari . During the last century, the field identification of A. hahneli , A. picta and other similar species has challenged biologists; with the description of A. imasmari sp. nov. there is another choice to distinguish from. The Quechua language is spoken by the Incan people indigenous to Andean Peru and the adjacent lower elevation areas where A. imasmari is known to occur.
Definition. A small species of cryptically-colored frog assigned to Ameerega due to the presence of maxillary and premaxillary teeth, the first finger being longer than the second, and basal webbing occurring between toes II- IV. The species can be characterized by the following combination of characters: (1) mean SVL of adult males 18.6 mm (range 18.3–19.0 mm), mean SVL of adult females 20.7 mm (range 19.8–21.8 mm) ( Table 1 View TABLE 1 ); (2) dorsal skin granular, especially on the back and the dorsal surfaces of the legs, dorsal surfaces of forelimbs lightly granular, flanks and venter non-granular; (3) finger III expanded in adults; (4) absence of lateral fringes and basal webbing on fingers; (5) maxillary and premaxillary teeth present; (6) fingers and toes discs expanded, larger in toes II, III, and IV; (7) basal webbing between toes II–III and III–IV; (8) finger I longer than finger II; (9) toe III length reaches or surpasses the middle of the central sub-articular tubercle of toe IV; (10) metatarsal fold absent; (11) tympanum conspicuous and small; (12) in life, base dorsal coloration of the head, back, and limbs are dark brown and finely mottled in brown; flanks black to dark brown; (13) oblique lateral stripe absent; (14) usually with white or cream dorsolateral stripes extending from loreal region to groin; (15) labial stripes present that match dorsolateral stripe coloration, starting behind nares and ending at forelimbs; (16) flanks black to dark brown, most individuals have bright white or cream, occasionally slate-blue, spotting or marbling extending from venter; (17) venter smooth, golden to light blue, with dense black marbling; (18) yellow to red spots present posterodorsally at the insertions of the forelimbs and hindlimbs, and on medial face of tibia; (19) limbs light to dark brown on dorsal surfaces, ventral surfaces of forelimbs are light blue distally, yellow proximally. Underside of head pigmented as the venter but often darker (i.e. more black marbling), especially in males; (20) iris dark brown with golden ring around pupil; (21) in preservative, all lighter pigments and flash colors fade to white or gray; (22) advertisement call is a short ‘peep’ repeated 2.1–2.6 times per second for several minutes; each note is short (mean of 0.17 s), with the dominant frequency from 4300–4500 Hz; (23) presence of exotrophic tadpoles carried to small terrestrial pools, where they are deposited in groups.
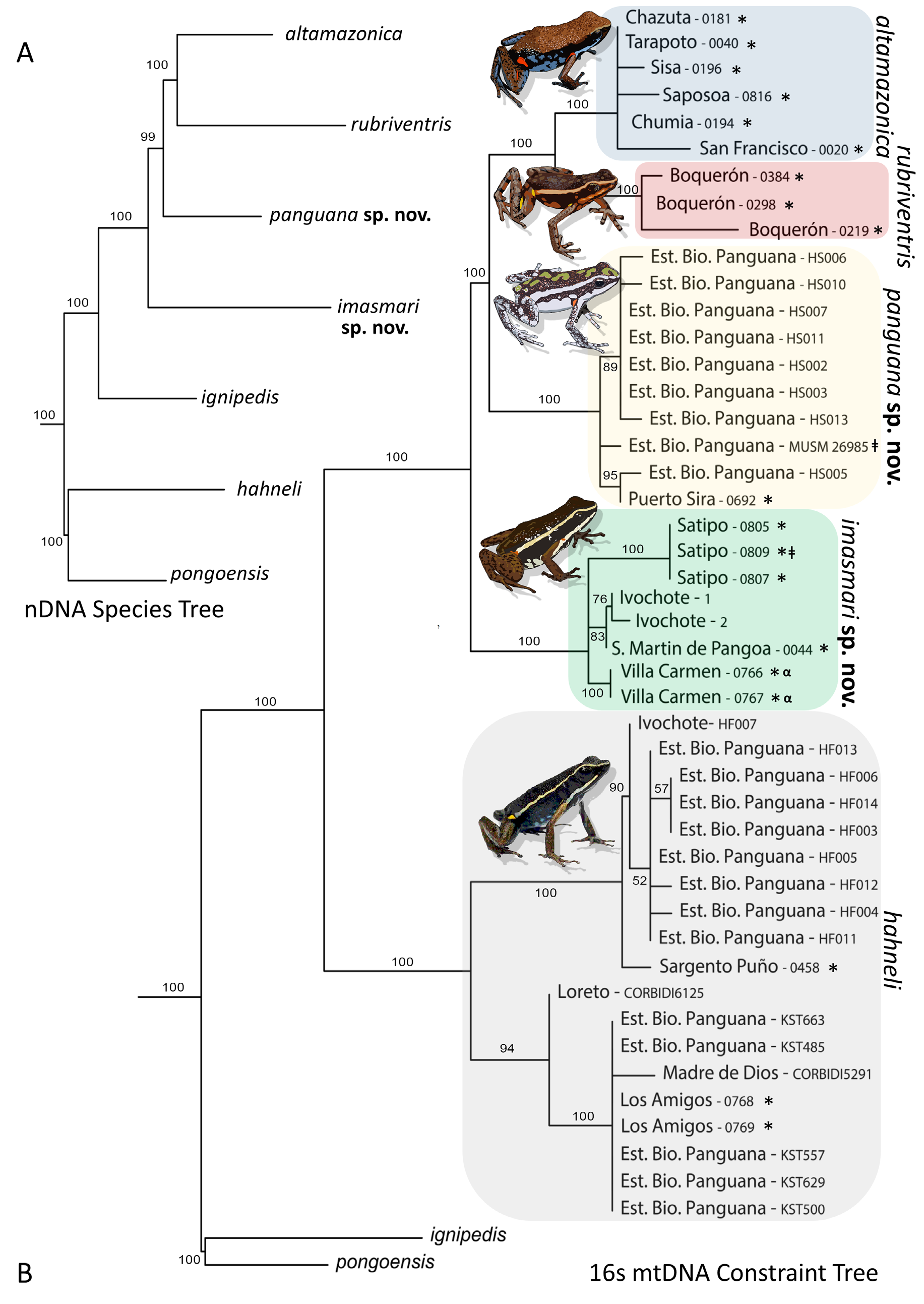
Diagnosis. Though they are not sister species ( Fig. 2 View FIGURE 2 ), southern populations of A. imasmari and northern populations of A. altamazonica are phenotypically similar and confidently distinguishing one from another may not be possible in the absence of genetic data ( Figs. 2 View FIGURE 2 & 3 View FIGURE 3 ). In A. hahneli sensu stricto, the width of each dorsolateral stripe decreases to a very fine stripe on the snout, occasionally disappearing (versus being wide and remaining the same width around the entire snout in A. imasmari ), and its advertisement call consists of 6–10 notes per second for several minutes (versus 2–3 notes per second in A. imasmari ). Another similar species is Ameerega panguana sp. nov., with some individuals morphologically similar, but with white dorsolateral stripes and venter coloration (versus golden in Satipo populations of A. imasmari ), and with an advertisement call consisting of 1–2 notes per second with a dominant frequency of 4.9–5.5 kHz (versus 2-3 notes per second with a dominant frequency of 4.3–4.5 kHz in A. imasmari , Table 3 View TABLE 3 ). Other species similar in appearance to A. imasmari include A. boliviana , A. picta sensu stricto, A. petersi , A. simulans , A. ignipedis , A. pulchripecta (all of which have yellowish or green dorsolateral stripes and blue venters), A. shihuemoy (pink dorsolateral stripes), and A. rubriventris (reddish-orange venter instead of a blue or golden venter in A. imasmari ).
Measurements of holotype (in mm). SVL 19.9; FL 9.8; TL 10.9; KK 19.5; FoL 9.4; HaL 6.0; HL 7.2; HW 6.8; BW 6.4; UEW 2.2; IOD 2.4; IND 2.5; TD 1.5; ED 2.4; DET 0.5; L1F 4.1; L2F 4.0; W3D 0.6; W3F 0.4. For paratypes see Table 2.
Description of adults. Little to no sexual dimorphism is apparent except for males being slightly smaller, possessing vocal slits and a subgular vocal sac. Teeth absent; tongue gray, ovoid, attaching anteriorly. Head widest at jaw articulations, slightly wider than body in most individuals (head width at tympanum 85.0–120.0% of body width at axillae); head width 26.8–32.8 % of SVL. Snout sloping laterally; bluntly rounded dorsally; truncate ventrally. Nares situated and directed posterolaterally to the tip of snout; nares visible from front and below but not from above. Canthus rostralis sloped, slightly rounded; loreal region nearly vertical and slightly concave. Interorbital distance nearly same width of superior upper eyelid. Eye large and prominent, with a maximum diameter of 7.9–8.9% of the snout vent length; pupil rounded and horizontally elliptical. Tympanum circular, partially concealed posterodorsally, lacking tympanic annulus; tympanum width 47.8–62.5% of eye diameter. Supratympanic fold absent.
Hands relatively small, length 23.7–29.6% of SVL. Relative length of appressed fingers: III> IV> II ≈ I. Discs moderately expanded, disc on finger III 1.4–2 times width of finger below disc. A large, circular outer metacarpal tubercle on median base of palm; a smaller inner metacarpal tubercle on base of finger I; one well developed and prominent subarticular tubercle on fingers I and II, two on fingers III and IV.
Hind limbs relatively small, femur 42.9–49.7% of SVL, tibia 42.9–53.0% of SVL. Relative lengths of appressed toes IV> III> V> II> I; first toe short, barely reaching bottom of subarticular tubercle on base of second toe, with unexpanded disc; toes II and III with barely expanded discs (much smaller than finger discs), and toes IV and V with discs expanded (disc 1.3–1.5 times broader than adjacent phalanx). Inner and a smaller outer metatarsal tubercle present, somewhat protuberant with rounded surfaces. One slightly protuberant subarticular tubercle present on toes I and II, two on toes III, IV, and V. Hands and feet lacking supernumerary tubercles, lateral fringes, and webbing. No toe fringes.
Vocalization. The advertisement call for A. imasmari ( Figs. 5 View FIGURE 5 & 6 View FIGURE 6 ) is a short ‘peep’ repeated 2.1–2.6 times per second for several minutes. Each note is short (mean of 0.17 s), with the dominant frequency from 4.3–4.5 kHz. This single-note advertisement call is given most frequently in the afternoon and evening as males chorus in small groups. We also observed a second call type in A. imasmari consisting of three to four notes, occasionally two notes in quick succession (separated by a mean of 13 ms, of silence), repeated once every 2-30 seconds. The latter call appears to function as an aggressive or territorial call and is most frequently heard in the early morning and early evening ( Schlüter 1980).
Distribution and Natural History. Ameerega imasmari is distributed throughout the east-Andean versant and surrounding lowlands of southern Peru, across the Fitzcarrald Arch, at elevations of ca. 200-400 m ( Fig 7 View FIGURE 7 ). This species appears to be sparsely distributed throughout regions Cuzco, Junín, and Pasco, and may extend into Madre de Dios and Ucayali. This species is locally abundant around Satipo (Pasco) and Villa Carmen (Cusco). Elsewhere it appears to be rare, or more likely undetected. This species is common and can be locally abundant in disturbed forested habitats (Purmas) nearby permanent water sources, which collectively provide refugia from the midday heat and habitat for reproduction. Ameerega imasmari does not appear to be commonly found in secondary and old-growth forests. This species commonly co-occurs with six other Ameerega species: A. trivittata , A. shihuemoy , A. simulans , A. picta , A. macero sensu stricto and A. hahneli sensu stricto.
Conservation status. Following the IUCN Red List criteria 3.1 (IUCN, 2012), we suggest A. imasmari be listed as Least Concern (LC) or Data Deficient (DD) under the following criteria: (1) we estimate its extent of occurrence at 41,690 km 2 (as pictured in Fig. 7 View FIGURE 7 ), and part of this range lies within two national parks (Otishi and Manu), a protected forest (Pui Pui), a natural sanctuary (Megantoni) and a communal reserve (Asháninka); (2) it occurs undisturbed habitat and disturbed areas; (3) population sizes are unknown, but assumed to be large given the abundance of forested habitats and tentative large range; (4) populations do not appear to be declining; and (5) demand for the pet trade is presumed to be low.






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Ameerega imasmari
| Brown, Jason L., Siu-Ting, Karen, May, Rudolf Von, Twomey, Evan, Guillory, Wilson X., Deutsch, Michael S. & Chávez, Germán 2019 |
Ameerega
| Siu Ting 2012 |
Epipedobates hahneli:
| Roberts 2006 |