
Anonchotaenia kornyushini, Mariaux & Georgiev, 2020
|
publication ID |
https://doi.org/ 10.5852/ejt.2020.616 |
|
publication LSID |
lsid:zoobank.org:pub:144F0449-7736-44A0-8D75-FA5B95A04E23 |
|
DOI |
https://doi.org/10.5281/zenodo.4332096 |
|
persistent identifier |
https://treatment.plazi.org/id/7CB90CCB-EC69-401F-AAA6-10A697956E1A |
|
taxon LSID |
lsid:zoobank.org:act:7CB90CCB-EC69-401F-AAA6-10A697956E1A |
|
treatment provided by |
Plazi |
|
scientific name |
Anonchotaenia kornyushini |
| status |
sp. nov. |
Anonchotaenia kornyushini sp. nov.
urn:lsid:zoobank.org:act:7CB90CCB-EC69-401F-AAA6-10A697956E1A
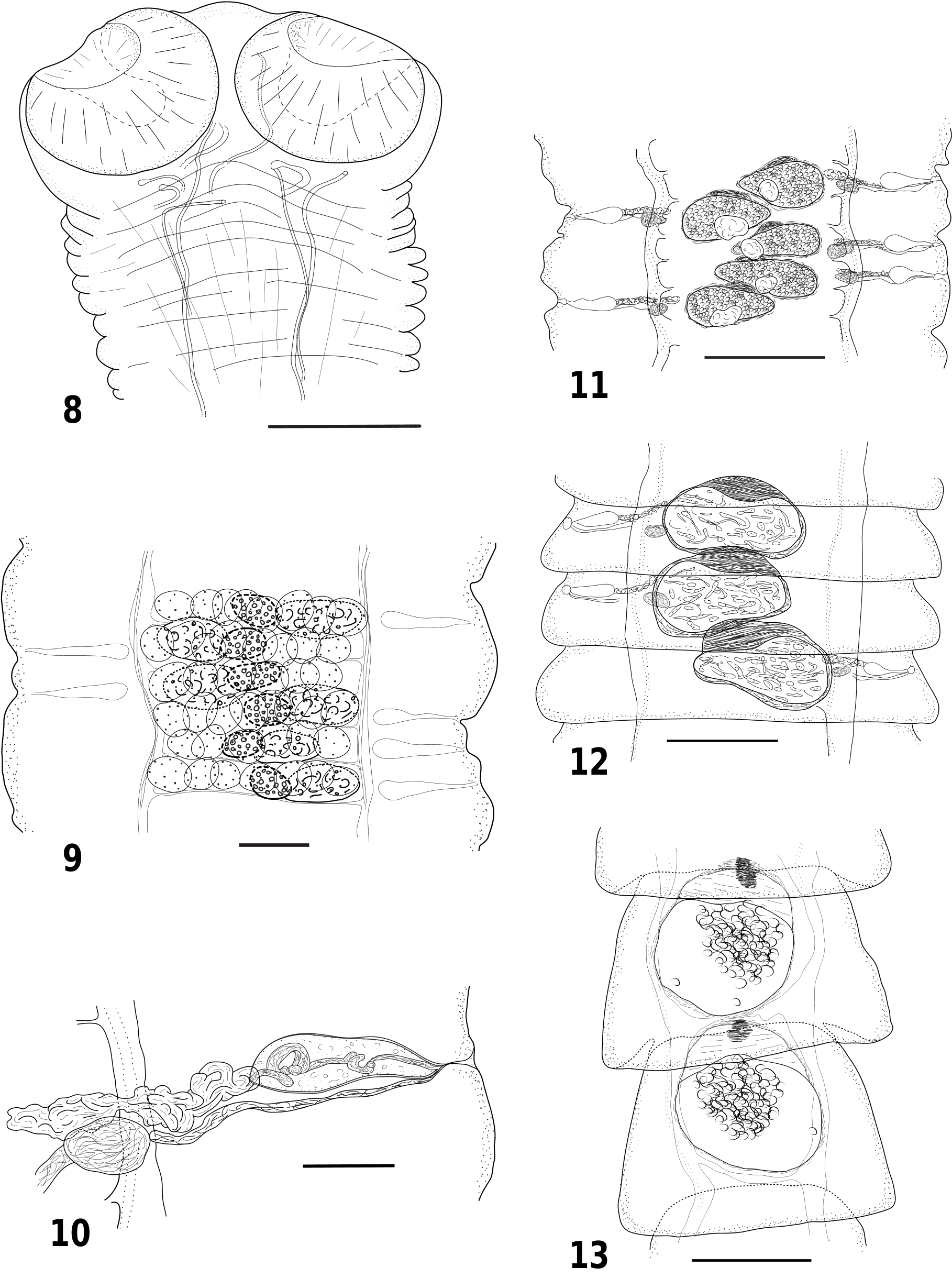
Figs 8–13 View Figs 8–13
Etymology
The species is dedicated to our colleague Prof. Vadim V. Kornyushin (Kiev, Ukraine), a dedicated cestode taxonomist with numerous contributions to the knowledge of the family Paruterinidae .
Material examined
Holotype
MALAYSIA • Selangor, University of Malaya’s Gombak Field Station ; 3.32° N, 101.77° E; 280–350 m a.s.l.; 9 Aug. 2010; MHNG-PLAT-121232. GoogleMaps
Paratypes
MALAYSIA • 2 specs; same collection data as for holotype; MHNG-PLAT- 121233 GoogleMaps .
Hologenophore
MHNG-PLAT- 121234 [field # MAL-075/C3]. Partial COI sequence, Genbank MN 590285 View Materials .
Type host
Trichastoma malaccense (Hartlaub, 1844) ( Passeriformes , Pellorneidae ).
Description
Body of medium size, 42 mm (n = 1) long, with maximum width 930 at level of early gravid proglottides; consisting of 540 proglottides. Mature proglottides acraspedote, about 15–20 times as wide as long; progressively becoming longer, 8–10 times wider at pregravid stage, then becoming markedly craspedote when gravid; last gravid proglottides almost as long as wide. Scolex rounded, 710–760 (730, n = 3) in diameter, poorly delineated from strobila. Suckers strong, muscular, 300–350 (327, n = 12) in diameter. Rostellum and rostellar sac absent ( Fig. 8 View Figs 8–13 ). Unsegmented zone 1.7–1.8 mm long behind posterior margin of suckers. Genital pores irregularly alternating in short series (mostly 1 or 2, maximum of 5 consecutive pores on the same side), opening in middle of lateral margin of proglottis. Genital ducts ventral to osmoregulatory canals. Ventral osmoregulatory canals up to 75 wide, connected posteriorly in each proglottis by transverse anastomosis. Dorsal osmoregulatory canals 5–10 wide. Genital atrium simple, with narrow infundibular orifice, 17–25 deep.
Testes large, spherical to slightly oval, 6–9 in number [6 (19%), 7 (37.5%), 8 (36%), 9 (7.5%)] (7.3, n = 53); 43–62 (50, n = 20) in diameter; arranged in single transverse row (with some partial overlapping) situated dorsally, occupying entire median field. Cirrus sac discrete, claviform, straight, with narrow poral half; not reaching osmoregulatory canals, 93–114 × 26–36 (103 × 30, n = 30). Cirrus unarmed. Internal vas deferens making a few loops in cirrus sac. External vas deferens coiled, situated antiporally to cirrus sac ( Figs 9–10 View Figs 8–13 ).
Vitellarium compact, oval, 45–55 × 58–80 (51 × 67, n = 20); aporal to ovary or partially overlapping its antiporal extremity dorsally. Ovary oval, 57–85 × 100–148 (68 × 123, n = 20); in poral half of median field. Mehlis’ gland not distinct. Vagina well-marked, opening posterior to male pore. Seminal receptacle oval to fusiform, on poral side of developing uterus. Uterus saccular, appearing centrally, dorsal to ovary in post-mature and pregravid proglottides, expanding laterally, becoming teardrop-shaped with thicker poral extremity ( Fig. 11 View Figs 8–13 ), then becoming more and more globular. Uterine wall thick. Paruterine organ consisting of fibrillar tissue, appearing rapidly adjacent to developing uterus, directed anteriorly to it and progressively becoming thicker ( Figs 12–13 View Figs 8–13 ). Eggs initially spherical, becoming vermiform in gravid proglottides, gradually filling uterus. At spherical stage, oncospheres 12.5–16.5 (14, n = 20) and eggs 18–20.5 (19.5, n = 20) in diameter. Embryonic hooks (poorly visible) ca 8 in length. Eggs passing into paruterine cavity not observed, but most developed proglottides with antero-central densification suggesting formation of this cavity.
Remarks
This material belongs to the genus Anonchotaenia Cohn, 1900 as defined by Mariaux (1991) or Georgiev & Kornyushin (1994). The most recent review of its diversity was by Phillips et al. (2014), who presented a summary of the main characters of the known species. Subsequently, a single additional species, A. adhiraji Banerjee, Manna & Sanyal, 2018 , was described in India ( Banerjee et al. 2018). This allows the easy comparison of our material with the 30 known species of Anonchotaenia . Most of Anonchotaenia spp. clearly differ by more than one of the characters listed by Phillips et al. (2014) (scolex and sucker diameters, position of genital ducts, testis number, cirrus sac extent and size) and only 2 species are similar to our material, i.e., A. castellanii Fuhrmann & Baer, 1943 , described from an Ethiopian laniid bird, and A. globata ( von Linstow, 1879) , initially described from a parid bird from Germany ( von Linstow 1879). The former species exhibits close similarities with our material; however, its testis number (9–10) is only marginally overlapping our observations (6–9, with a dominant proportion of 7–8 testes). Anonchotaenia castellanii further differs from our material in showing a much more muscular cirrus-sac as well as a strong atrial sphincter ( Fuhrmann & Baer 1943). Furthermore, Eurocephalus rueppellii Bonaparte, 1853 , the host of A. castellanii , is endemic to Eastern Africa.
Anonchotaenia globata has been reported from a wide range of Passeriformes all over the world ( Matevosyan 1969). It also resembles our material for most characters, even if a precise description of this species is difficult to infer from the multiple reports of this taxon. Kornyushin (1989) believed that the reports of A. globata should be referred to a complex of species. Philips et al. (2014) presented metrical data based on summarising known ranges reported for A. globata by many authors from various hosts and various geographical origins (but erroneously mentioning in their table that these data come from the original description only). Even considered in this wide and doubtful taxonomic context, A. globata is characterized by a smaller number of testes (4–5) than our material, though a larger range has occasionally been reported for this species. Although a few other unusual morphological characters (such as the changes of the shape of the paruterine organ during its development) seem promising for specific identifications in the group ( Phillips et al. 2014), they still lack a complete comparative basis. Testis number presently remains crucial, and the most reliable criterion to identify Anonchotaenia spp. at the morphological level ( Mariaux 1991). We consider it sufficient to justify the placement of our material in a new species that we name Anonchotaenia kornyushini sp. nov.
| MN |
Museu Nacional, Universidade Federal do Rio de Janeiro |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |