
Calliaxina chalmasii ( Brocchi, 1883 ) Brocchi, 1883
|
publication ID |
https://doi.org/ 10.5281/zenodo.210053 |
|
publication LSID |
lsid:zoobank.org:pub:1AF0BD07-97AB-4018-B973-98AB67951238 |
|
DOI |
https://doi.org/10.5281/zenodo.5662220 |
|
persistent identifier |
https://treatment.plazi.org/id/BF006C37-FF88-7D42-FF61-A25D3240FECD |
|
treatment provided by |
Plazi |
|
scientific name |
Calliaxina chalmasii ( Brocchi, 1883 ) |
| status |
comb. nov. |
Calliaxina chalmasii ( Brocchi, 1883) comb. nov.
( Figs. 3 View FIGURE 3 A–B, 4A–R, 5A–E, 6A–E)
Callianassa Chalmasii Brocchi, 1883: 6 View in CoL , 7, pl. 5, figs. 7, 8.
Callianassa Chalmasi. View in CoL — Glaessner, 1929: 77.
Callianassa chalmasii . — Müller, 1979: 274, pl. 1, fig. 5; Schweitzer et al., 2010: 34.
Calianassa chalmasii .— Lőrenthey, 1897: 150, 160, 168; Lőrenthey, 1898 a: 93, 105, 114; Lőrenthey, 1898 b: 9, 129, 130, 155; Lőrenthey, 1898 c: 102; Lőrenthey, 1904: 161; Lőrenthey, 1913: 332; Lőrenthey in Lőrenthey & Beurlen, 1929: 33, 65, pl. 2, figs. 15a, 15b.
‘ Callianassa View in CoL ’ chalmasii .— Müller, 1984: 50, pl. 2, figs. 3–7.
Callianassa espichelensis Veiga Ferreira, 1961: 479 –481, pl. 1, figs. 1–11; Schweitzer et al., 2010: 34.
Calianassa rákosiensis Lőrenthey, 1897: 160 , 161, 168, 169 (new synonym); Lőrenthey, 1898 a: 105, 114, 115; Lőrenthey, 1898 b: 131, 132, pl. 9, fig. 4; Lőrenthey, 1898 c: 103, 104, pl. 9, fig. 4;
Calianassa rákosensis .—Lőrenthey in Lőrenthey & Beurlen, 1929: 33, 66, 67, pl. 2, fig. 14.
Callianassa rákosensis .— Müller, 1979: 274, 276, pl. 1, fig. 4.
Callianassa rakosiensis .— Glaessner, 1929: 89; Schweitzer et al., 2010: 36.
‘ Callianassa View in CoL ’ rakosiensis .— Müller, 1984: 51, pl. 3, figs. 1, 2.
Callianassa cf. rákosensis .—Bachmayer, 1953: 242; Bachmayer & Tollmann, 1953: 312.
?„ Callianassa View in CoL “ sp. 2— Hyžný, 2011: 42, fig. 4E, F.
non Callianassa chalmasii . — De Angeli et al., 2009: 168, figs. 2C, D; Garassino et al., 2012: 18. (= Calliaxina punica )
Diagnosis. Calliaxina with both P1 propodi quadrate, upper and lower margins keeled; fixed finger of major propodus with diagonally oriented ridge, occlusal margin toothed; minor propodus with well developed tuberculated depression on the base of fixed finger.
Description. First pereiopods (chelipeds) with major and minor cheliped strongly developed, subequal in size, similar, laterally compressed.
Carpus unarmed, slightly higher than long, triangular in shape, upper and lower margin keeled, terminating in blunt corner at articulation with propodus, lower margin together with proximal margin forming a single rounded edge.
Both major and minor propodus quadrate, about as high as long, sometimes longer or shorter than high, converging distally; upper and lower margins keeled, terminating in blunt corners at articulation with carpus; keel on the upper margin forming sharp extension distally; keel on the lower margin distinctly broader proximally, forming flat edge; inner surface of propodi smooth with distinct setal pores; setal pores on the upper margin large, elongated in vertical aspect, i.e. perpendicular to the longitudinal axis of the limb.
In major propodus (previously described as Callianassa chalmasii ), area at the articulation with dactylus covered with fine tubercles; fixed finger short, triangular, with distinct obliquely oriented ridge on the lateral surface; occlusal margin of fixed finger with distinct tooth followed by serrated edge; lower margin of propodus slightly convex proximally.
In minor propodus (previously described as C. rakosiensis ), area at the articulation with dactylus distinctly depressed, covered with tubercles; depression forming sharp edge at its lower margin, which is parallel to the lower margin of propodus distally and distinctly rounded proximally; lower margin of propodus usually distinctly straight.
Dactylus slightly curved, unarmed.
Variations. The material shows a degree of variation in certain characters present on propodus. The length/ height ratio is variable; both forms with distinctly longer ( Figs. 4 View FIGURE 4 E, I) or shorter propodus ( Fig. 4 View FIGURE 4 K) are present, although the majority of material exhibits more-less equal ratio (e.i. quadrate manus) ( Table 2 View TABLE 2 ). Short morphotype has already been depicted, but not commented upon by Müller (1984: pl. 2, fig. 3).
Lower margin of minor propodus is sometimes concave at its distal end ( Figs. 4 View FIGURE 4 M, P). In such cases the fixed finger bears a distinct tooth on its occlusal margin.
Material examined. Holotype of Callianassa rakosiensis representing right propodus (FI 29); Müller (1984: 51) stated that it had been probably lost, however, it was found during a visit of the Hungarian Geological Institute. The holotype of C. chalmasii has not been found and at present it should be considered to be lost as already stated by Müller (1984: 51). Material from Rákos collected by P. Müller and deposited at the Hungarian Natural History Museum comprises the right propodus of C. rakosiensis (M.86.244, illustrated also by Müller 1984: pl. 3, figs 1–2.) and several isolated propodi of C. chalmasii (M.86.275). Newer material collected by P. Müller is deposited under numbers PAL 2011.29, PAL 2011.32–37 (locality Rákos), PAL 2011.30 (Örs vezér tere), and PAL 2011.31 (Gyakorló út) ( Fig. 4 View FIGURE 4 ). Material from Rákos collected by the author is deposited under numbers KGP-MH RA020–037 ( Fig. 4 View FIGURE 4 ). Additional material comprises five specimens from the Fenk quarry (NHMW-2011-0167- 0116–0120) ( Fig. 5 View FIGURE 5 ) and several propodal fragments (one articulated with carpus) from Dúbravská hlavica (SNM- Z 37570, 37571); KGP-MH DH074–076) ( Fig. 6 View FIGURE 6 ).
Measurements and details about the specimens are summarized in Table 2 View TABLE 2 .
Other material. For comparative purposes, the extant material of several Eucalliacinae taxa have been examined (all deposited in NHMW), namely Calliaxina novaebritanniae ( NHMW 25399, 25400); Eucalliax aequimana ( NHMW 19365); and E. kensleyi ( NHMW 16779).
Occurrence. The oldest occurrence of Calliaxina chalmasii comb. nov., as reported herein, is that from the Fenk quarry, Gross Höflein, Austria, which is middle 'Badenian' (upper Langhian) in age. From the upper 'Badenian' (lower Serravallian), the species is known from the surroundings of the Budapest area, Hungary ( Müller 1984). It was reported also from the Middle Miocene of Portugal ( Veiga Ferreira 1961). Thus, it appears that during Miocene the species was widespread in the Central Paratethys. Since, the Late Miocene, when the seaways between these two palaeogeographic areas were closed and the Central Paratethys became Lake Pannon ( Rögl 1998, 1999; Harzhauser et al. 2002; Harzhauser & Piller 2007), the species might have migrated into the Mediterranean where it gave rise to extant Calliaxina punica . Following the chronological approach (see below) the material of Callianassa chalmasii reported from the Early Pliocene of Italy ( De Angeli et al. 2009; Garassino et al. 2012) may in fact represent C. punica , as numerous extant decapod species are known from their Pliocene occurrences. This conclusion may be supported by the reported co-occurrence of presumed C. chalmasii and Calliaxina cf. C. punica at one of the Italian localities ( Garassino et al. 2012).
Remarks. In description of Callianassa rakosiensis from the Middle Miocene of Hungary Müller (1984: 51) stated: “The species is remarkably similar to C. chalmasii in its general form, size and the distribution of pores. The form of the ridge on the finger and the position of the tooth is completely different in the two species and no trend was observed toward a transitional form. Thus their independence seems highly probable though a close relation is likely.” What Müller (1984) recognized as two separate species in fact represents minor ( C. rakosiensis ) and major chelae ( C. chalmasii ) of a single species. Müller's observation of no transitional form and simultaneously the presence of both morphotypes at the same localities (Rákos, Gyakorlo út and Örs vezér tere) support the synonymisation proposed here.
Callianassa espichelensis described by Veiga Ferreira (1961) has already been considered as a junior synonym of C. chalmasii ( Müller 1984) . Figures ( Veiga Ferreira 1961: fig. 1) clearly show the same morphotype. Most illustrated specimens represent major chelae ( C. chalmasii morphotype); however, one specimen (fig. 1.4) seems to represent the minor chela ( C. rakosiensis morphotype). It is illustrated from the inner side, so, the depression present on the outer lateral surface is not visible. Veiga Ferreira (1961: 480), however, mentioned the presence of a ridge, depression, and also faint tuberculation at the base of the fixed finger. I concur with Müller (1984), contrary to Schweitzer et al. (2010), and consider C. espichelensis as a junior subjective synonym of C. chalmasii .
Hyžný (2011: 42) reported a burrow structure with the cheliped remains of two individuals identified as “ Callianassa View in CoL ” sp. 2 from the Gyakorló út locality. Although the material is insufficiently preserved, one individual clearly possesses equal or subequal chelipeds. Thus, it might represent C. chalmasii comb. nov., which has already been reported from that locality ( Müller 1984; this paper). The material itself is, however, fragmentary and poorly preserved. As taxonomically important characters are discussed herein are not readily visible, I am hesitant to consider it to be conspecific with C. chalmasii comb. nov.
Calliaxina chalmasii comb. nov. can be easily distinguished from all other Middle Miocene callianassid forms (for a review see Hyžný 2011: table 2); none of them has a well developed tuberculated depression on minor propodus at the base of the fixed finger. Moreover, C. chalmasii comb. nov. has laterally compressed chelipeds, a character that is uncommon in callianassid taxa co-occurring at the same localities. On the other hand, C. chalmasii comb. nov. is virtually indistinguishable from the extant forms, notably C. punica and C. novaebritanniae , on the basis of propodus and carpus alone. Characters on merus might resolve this issue. I concur with Müller (1984: 48) that: “identification of 12–15 milion year old species with extant ones should be done with caution”. Thus, a chronological approach (which operates with chronospecies rather than morphospecies) is followed here (see also Klaus & Gross 2010) and Calliaxina chalmasii comb. nov. is considered a separate species distinct from all extant congeners.
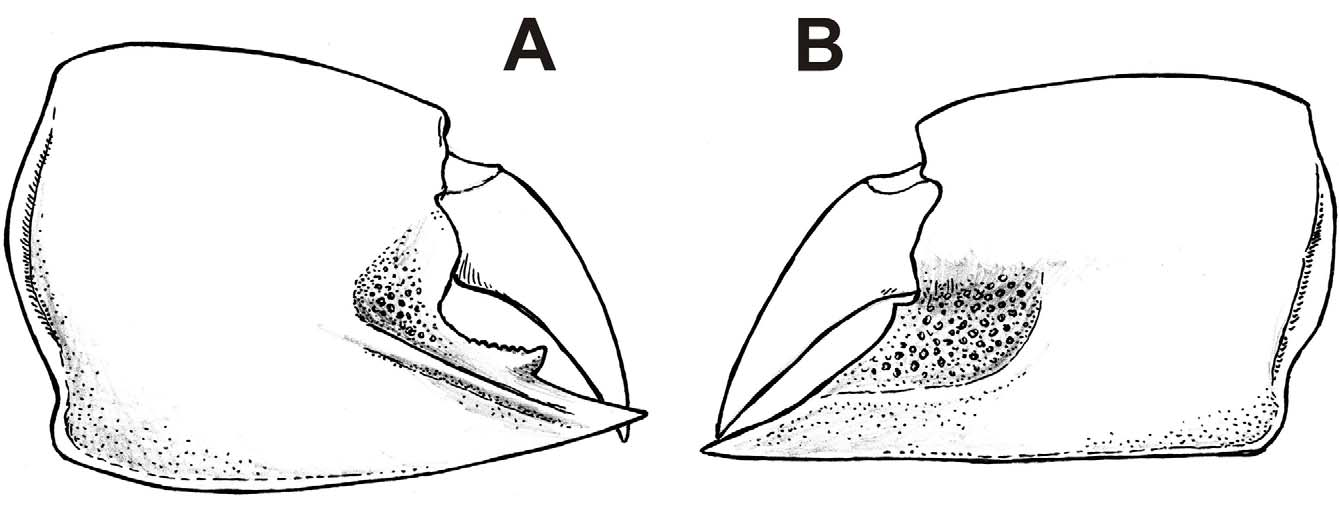



TABLE 2. Measurements (mm) of Calliaxina chalmasii (Brocchi, 1883) comb. nov. Abbreviations: M = major, m = minor,? = unknown; handedness: L = left; R = right; max. L = maximum length of the element, max. H. = maximum height of the element; measurements: ~ = approximate value.
| Specimen | M/m | handedness | element | max.L | max.H |
|---|---|---|---|---|---|
| KGP-MH DH-074 KGP-MH DH-075 KGP-MH RA-020 KGP-MH RA-021 KGP-MH RA-022 KGP-MH RA-023 | M M M M M M | L L L L L R | propodus propodus propodus propodus propodus propodus | 8.0 6.2 10.0 ~7.5 6.0 4.4 | 7.1 6.7 8.5 7.3 5.5 3.6 |
| KGP-MH RA-024 KGP-MH RA-025 KGP-MH RA-026 KGP-MH RA-027 KGP-MH RA-028 KGP-MH RA-029 | M M M m?? | L R R R R L | propodus propodus propodus propodus carpus propodus | 6.0 4.0 4.6 7.2 5.3 ~6.0 | 4.3 3.4 4.5 7.0 5.8 ~6.3 |
| KGP-MH RA-030 KGP-MH RA-031 | ?? | R R | carpus carpus | 5.0 5.5 | 5.7 6.0 |
| KGP-MH RA-032 KGP-MH RA-033 KGP-MH RA-034 KGP-MH RA-035 | ? M M m | L R L L | carpus propodus propodus propodus | 3.2 6.0 5.6 7.6 | 3.7 6.3 5.7 7.7 |
| KGP-MH RA-036 KGP-MH RA-037 NHMW-2011-0167-0116 NHMW-2011-0167-0117 NHMW-2011-0167-0118 NHMW-2011-0167-0120a | m m? m m M | L R R R L R | propodus propodus carpus propodus propodus propodus | 5.0 6.8 5.7 5.1 7.3 7.0 | 6.0 6.2 5.6 5.0 7.3 6.7 |
| NHMW-2011-0167-0120b PAL 2011.29 PAL 2011.30 PAL 2011.31 PAL 2011.32 PAL 2011.33 | M m m m m M | L L L L L L | propodus propodus propodus propodus propodus propodus | 3.2 6.3 ~4.5 8.6 5.0 5.6 | 3.0 6.4 4.2 8.3 4.5 5.4 |
| PAL 2011.34 PAL 2011.35 PAL 2011.36 PAL 2011.37 SNM Z-37570 SNM Z-37570 | M M M M m m | R R R L R R | propodus propodus propodus propodus propodus carpus | 6.5 6.4 7.0 6.4 6.6 4.5 | 7.0 7.0 7.4 5.7 6.4 6.3 |
| SNM Z-375701 | M | L | propodus | 8.5 | 8.8 |
| NHMW |
Naturhistorisches Museum, Wien |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Calliaxina chalmasii ( Brocchi, 1883 )
| Hyžný, Matúš 2012 |
Callianassa chalmasii
| Garassino 2012: 18 |
| De 2009: 168 |
Callianassa
| Muller 1984: 50 |
Callianassa
| Muller 1984: 51 |
Callianassa chalmasii
| Schweitzer 2010: 34 |
| Muller 1979: 274 |
Callianassa rákosensis
| Muller 1979: 274 |
Callianassa espichelensis
| Schweitzer 2010: 34 |
| Veiga 1961: 479 |
Callianassa
| Glaessner 1929: 77 |
Calianassa rákosensis
| Lorenthey 1929: 33 |
Callianassa rakosiensis
| Schweitzer 2010: 36 |
| Glaessner 1929: 89 |
Calianassa chalmasii
| Lorenthey 1929: 33 |
| Lorenthey 1898: 93 |
| Lorenthey 1898: 9 |
| Lorenthey 1898: 102 |
| Lorenthey 1897: 150 |
Calianassa rákosiensis Lőrenthey, 1897 : 160
| Lorenthey 1898: 105 |
| Lorenthey 1898: 131 |
| Lorenthey 1898: 103 |
| Lorenthey 1897: 160 |
Callianassa Chalmasii Brocchi, 1883 : 6
| Brocchi 1883: 6 |