
Pinacocoleps tesselatus (Kahl, 1930) Foissner et al., 2008
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3637.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:E52CB032-BE68-4D07-8B8B-DB8F576D032F |
|
DOI |
https://doi.org/10.5281/zenodo.5631069 |
|
persistent identifier |
https://treatment.plazi.org/id/03CC87EC-0A37-CE23-FF12-D996645BA3EF |
|
treatment provided by |
Plazi |
|
scientific name |
Pinacocoleps tesselatus (Kahl, 1930) Foissner et al., 2008 |
| status |
|
Pinacocoleps tesselatus (Kahl, 1930) Foissner et al., 2008
( Figs 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 ; Tables 1 View TABLE 1 , 2 View TABLE 2 )
Basionym: Coleps tesselatus Kahl, 1930
Since no data based on silver impregnations are available, we suggest an improved diagnosis here according to investigations on both current and previous populations.
Improved diagnosis: About 60–85 μm in length in vivo, ovular shaped body; 21–25 ciliary rows, each composed of about 11 monokinetids and two perioral dikinetids; bar-shaped extrusomes mainly distributed along the oral basket; one terminal contractile vacuole; one macronucleus and one micronucleus; anterior and posterior main plates each with three teeth, respectively. Marine or brackish water.
Description of the Ningbo population: Body about 75–85 μm × 40–45 μm in vivo, shape rather constant and long oval-shaped, ratio of length to width about 2:1, usually slightly narrowed in mid-body where the main armour plates abut, circular in cross-section. Anterior end transversely truncate and crown-like due to the pointed ends of the secondary tier plates; posterior end moderately rounded ( Figs. 1 View FIGURE 1 A; 2A). One large, globular to slightly ellipsoidal macronucleus usually near mid-body, with an average length: width ratio of 1.1, about 15 μm across in vivo; one closely associated micronucleus about 4 μm across in size ( Figs. 1 View FIGURE 1 A, D; 2J). One terminal contractile vacuole, about 10–15 μm in diameter, directed canal and excretory pore not detected. Extrusomes bar-shaped, located mainly in the oral opening, invisible in vivo because of their small size (ca. 4 × 0.3 μm), darkly stained in the silver carbonate impregnated specimens ( Fig. 2 View FIGURE 2 H). Cytoplasm colorless, usually containing several food vacuoles (10 μm in diameter) and many granules (2–3 μm across). Swimming moderately fast in a counterclockwise helical path.
Armour with Pinacocoleps - type plate ( incurvus - type in Foissner et al. 2008), composed of six tiers: circumoral tier, anterior secondary tier, anterior main tier, posterior main tier, posterior secondary tier, and caudal tier, each composing an average of 22 rectangular plates. Circumoral tiers hardly recognizable in vivo. Anterior secondary plates approximately triangular shape, about 15 μm in length, with acutely pointed anterior ends forming a crown-like pattern, each with three teeth and including two ciliary outlets, one or two of them with anterior spines ( Figs. 1 View FIGURE 1 B; 2B). Anterior main plates rectangular shape, approximately 25 μm long, usually with three conspicuous teeth, including four ciliary outlets ( Figs. 1 View FIGURE 1 B; 2C). Posterior main plates rectangular shape, approximately 23 μm long, with three conspicuous teeth, and including three ciliary outlets ( Figs. 1 View FIGURE 1 B; 2D). Posterior secondary plates approximately trapezoidal, approximately 13 μm long, with single teeth, and including two ciliary outlets, rarely with sturdy posterior spines ( Figs. 1 View FIGURE 1 B; 2E). Caudal tier visible only in oblique or posterior polar views, with posterior spines up to 8 μm long ( Figs. 1 View FIGURE 1 B; 2G). Fine structure of armour plates as shown in Figs. 1 View FIGURE 1 B; 2B–F: inconspicuous ovate windows near right plate margin; a serious of teeth on right edge arranged alternately with windows, connected with thickened longitudinal ridge by dominant bridges; left margin very smooth and slightly curved.
Oral opening occupying central region of anterior pole. Circumoral kinety composed of paired basal bodies, interrupted at site adoral organelles ( Fig. 1 View FIGURE 1 C). Three slightly obliquely arranged adoral organelles: organelles 1 and 2 each consisting of four pairs of kinetosomes; organelle 3 consisting of three pairs of kinetosomes ( Figs. 1 View FIGURE 1 C; 2I). Oral basket inconspicuous in vivo, distinct in silver carbonate impregnated specimens, and about 20 μm long ( Fig. 2 View FIGURE 2 H).
Somatic cilia about 8 μm long and very regularly arranged, forming an average of 11 transverse circles and 22 longitudinal rows ( Figs. 1 View FIGURE 1 D; 2G and Table 1 View TABLE 1 ). Anterior end of longitudinal ciliary rows with two dikinetids each, forming the perioral ciliature. Perioral portion curved rightwards underneath the adoral organelles ( Figs. 1 View FIGURE 1 C; 2I). One caudal cilium about 25–30 μm long.
Abbreviations: CV = coefficient of variation in %, Max = maximum, Mean = arithmetic mean, Median = median value, Min = minimum, n = number of individuals examined, SD = standard deviation, SE = standard error of arithmetic mean.
Phylogenetic analyses: The SSU rDNA sequence is 1,641 bp long and available under Accession number KC349950 View Materials of GenBank database. In our preliminary analyses, the Prorodontidae is sister to the clade containing colepids and environmental samples. To further evaluate the phylogenetic position of P. tesselatus in the family Colepidae , we selected two Prorodon species as outgroups. Two clearly separated clades were recovered in both ML and BI methods: 1) P. tesselatus + Tiarina + Nolandia sinica + Apocoleps (ML 85%, BI 0.94); 2) Levicoleps + Nolandia nolandi + Coleps (ML 96%, BI 1.00). Species in Clade I are marine or brackish organisms, while the Clade II are comprised of freshwater species with one exceptional sequence from uncultured marine sample (GenBank accession number EU446396). None of the undescribed environmental sequences groups closely with P. tesselatus . Unfortunately, we couldn’t evaluate the genetic divergence within the genus Pinacocoleps as this is the first study in Pinacocoleps described with molecular data. Thus, further molecular studies are required to provide more data to compare with its congeners and uncover more reliable phylogenies of colepids.
Remarks and comparison: This species was first reported by Kahl (1930) from a marine and muddy moat in the Sylt Island, Germany with a brief description of the organism in vivo: 60–70 μm long, barrel-shaped body; anterior and posterior plates with three teeth; about 22–25 ciliary rows; three posterior spines ( Fig. 1 View FIGURE 1 E; Table 2 View TABLE 2 ). The organism we isolated matches the descriptions by Kahl (1930); thus, its identity is not in doubt.
Kahl provided a brief and significant description of this species in vivo, but the infraciliature has never been investigated. Furthermore, while there is no type available, the specimens of the Ningbo population of are deposited as vouchers.
Borror (1972) described a form under the name of Coleps tesselatus , which had an ovate to cylindrical body shape, 46–49 μm long after fixation in osmic acid fumes, anterior main plate with five teeth, and posterior main plate with three teeth, about 18 ciliary rows; three posterior spines ( Fig. 1 View FIGURE 1 G; Table 2 View TABLE 2 ). This isolate is very similar to Pinacocoleps similis (Kahl, 1933) , thus it is likely that Borror (1972) actually observed a population of the latter species rather than Pinacocoleps tesselatus (Kahl, 1930) .
Foissner et al. (2008) resurrected the genus Pinacocoleps created by Diesing (1865), and provided an amended diagnosis: Colepidae with spiny armour composed of six tiers with plates of the incurvus- type, number of adoral organelles not known. So far seven nominal species have been assigned to this genus: Pinacocoleps incurvus (Ehrenberg, 1833) Foissner et al., 2008 , P. tesselatus (Khal, 1930) Foissner et al., 2008 , P. pulcher (Spiegel, 1926) Foissner et al., 2008 , P. s i m i l i s (Kahl, 1933) Chen et al., 2010, P. heteracanthus (Noland, 1937) Chen et al., 2010 , P. s p i r a l i s (Noland, 1937) Chen et al., 2010, and P. arenarius (Bock, 1952) Chen et al., 2010 .
Data for taxon studied in the present paper are shown in bold. * = likely a misidentification (see text); ** = Specimens fixed 30 sec in osmic acid fumes; *** = Data counted from drawing;? = Data not available. AMP = anterior main plate; AS = anterior spine; PMP = posterior main plate; PS = posterior spine.
Among these species, Pinacocoleps arenarius (Bock, 1952) is very similar to P. tesselatus (Kahl, 1930) in terms of body length (70–75 μm vs. 75–85 μm) and its plate characters. Both have very weak ovate windows, anterior and posterior main plates with three teeth respectively. The fine structure of the armour plates and morphological characters of P. arenarius were insufficiently described, but it can be separated from P. tesselatus by the following features: body shape (rectangular, and laterally flattened vs. long oval-shaped, circular in crosssection), the number of posterior spines (10 vs. 3) ( Fig. 1 View FIGURE 1 I; Table 2 View TABLE 2 ; Bock 1952).
Morphologically, Pinacocoleps tesselatus is similar to P. s i m i l i s and P. i n c u r v u s. It is distinguished from P. similis by the structure of its armour plates: 1) each anterior secondary plate with single acutely pointed anterior end (vs. with two sharp anterior ends in P. s i m i l i s); 2) anterior main plate with three teeth and four ciliary outlets (vs. five teeth and six ciliary outlets in P. similis ); 3) a thick bridge connecting teeth to ridge present (vs. absent in P. similis ) and; three posterior spines (vs. at least five in P. similis ) ( Fig. 1 View FIGURE 1 F; Table2 View TABLE 2 ; Kahl 1933; Chen et al. 2010). Pinacocoleps tesselatus distinguished from P. i n c u r v u s, bybody shape (long oval-shaped vs. curved cylindrical); the number of anterior and posterior main plate teeth respectively (3, 3 vs. 7, 7); the number of ciliary rows (21–23 vs. 10–11); and the number of anterior spines (2 vs. 5) ( Fig. 1 View FIGURE 1 K; Table 2 View TABLE 2 ; Kahl 1930).
Pinacocoleps tesselatus also resembles P. p u l c h e r in the long oval-shaped outline, the body length (60–90 μm vs. ca. 100 μm), and the number of ciliary rows (21–25 vs. 20–22). However, Pinacocoleps tesselatus can be separated from P. pulcher by the structure of the armour plate: 1) thicken bridge connecting teeth to ridge present (vs. absent in P. p u l c h e r); 2) shape of left plate margin (very smooth and slightly curved vs. slightly serrated); 3) the number of main plate teeth (3 vs. 10); 4) the number of anterior and posterior spines respectively (2, 3 vs. 6, 7) ( Fig. 1 View FIGURE 1 L; Table 2 View TABLE 2 ; Kahl 1930; Spiegel 1926).
Noland (1937) described two species, namely Pinacocoleps heteracanthus , and P. spiralis , which resemble P. tesselatus in having Pinacocoleps - type plate, long oval or barrel-shaped body, and about 22–23 ciliary rows. However, these two can be clearly distinguished from P. tesselatus by the number of main plates teeth (9–11, 7– 9 in P. heteracanthus ; 7–8, 5– 6 in P. spiralis ; 3, 3 in P. tesselatus ). In addition, P. spiralis can be easily distinguished from other congeners by the longitudinal rows of plates arranged in a definitely spiral torsion ( Figs. 1 View FIGURE 1 H, J; Table 2 View TABLE 2 ; Noland 1937).
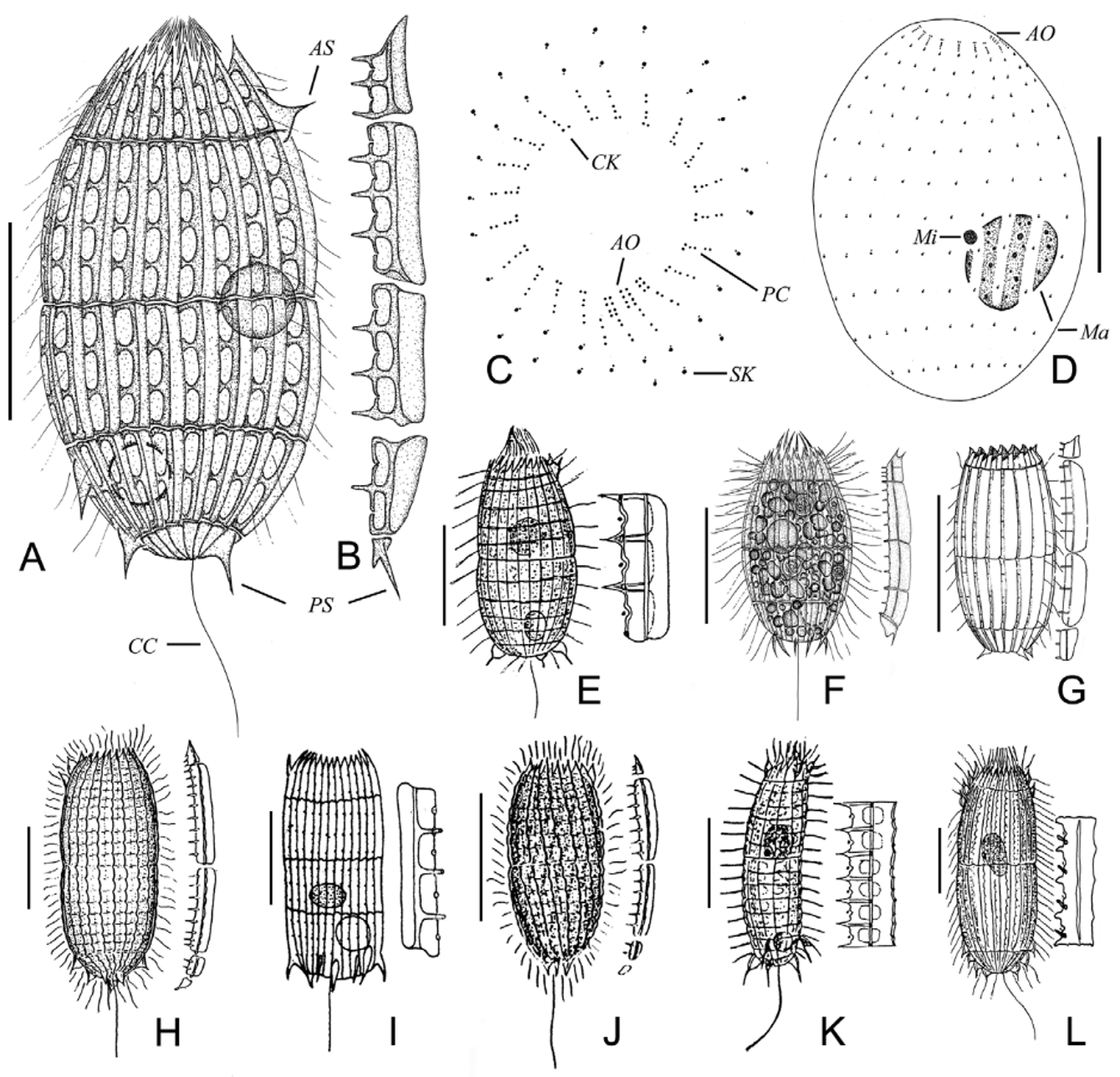
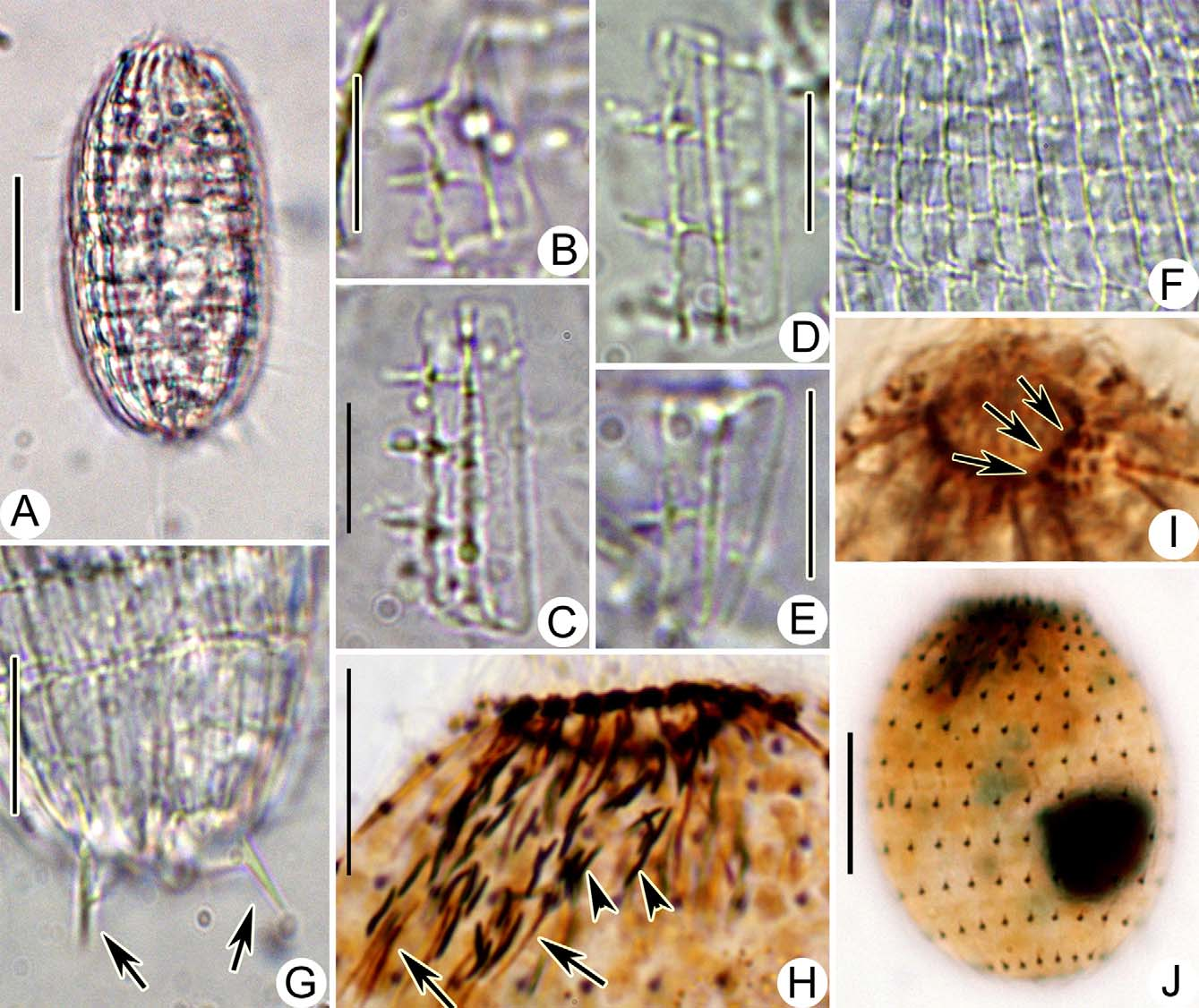
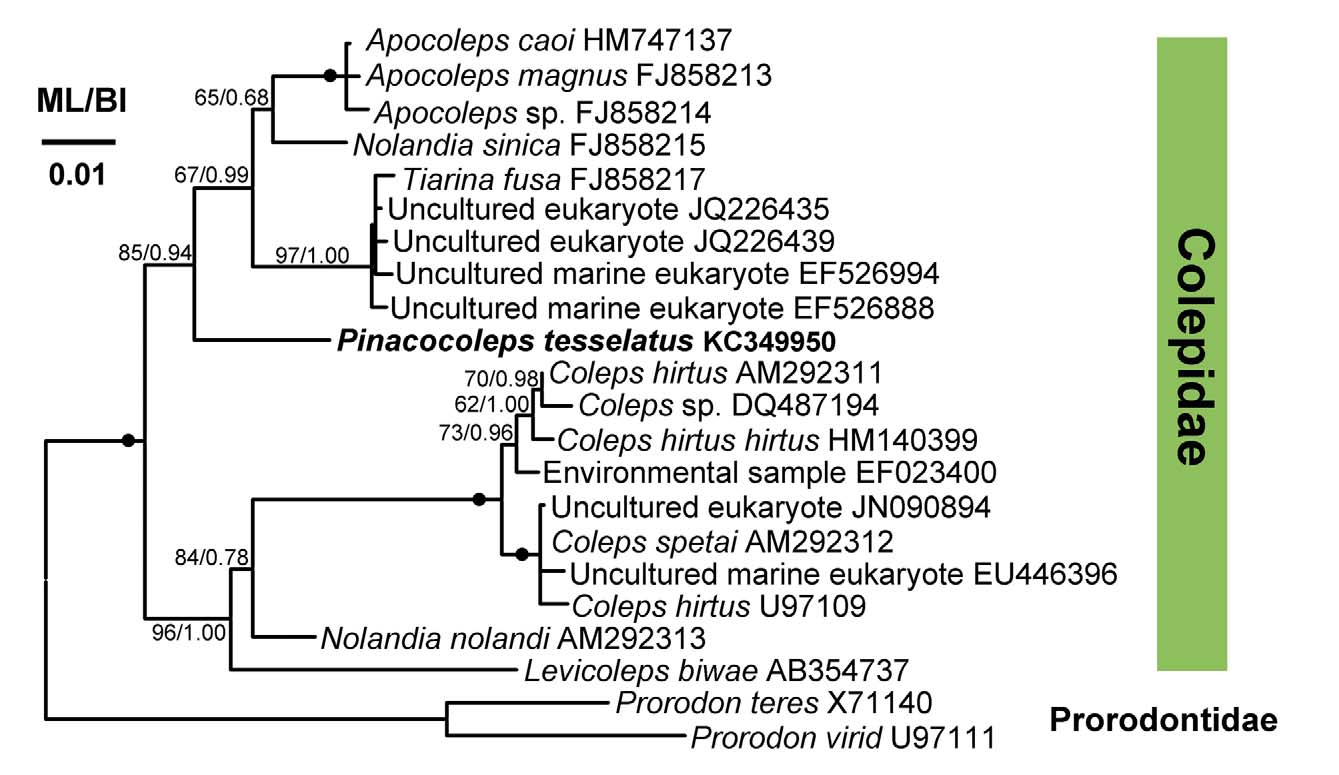
TABLE 1. Morphometric characteristics of Pinacocoleps tesselatus (Kahl, 1930) from silver carbonate impregnated specimens. All measurements in μm.
| Characters | Min | Max | Median | Mean | SD | SE | CV | n |
|---|---|---|---|---|---|---|---|---|
| Body length | 73 | 83 | 77.5 | 78.0 | 3.3 | 0.7 | 4.2 | 20 |
| Body width | 61 | 68 | 63.5 | 64.3 | 2.3 | 0.5 | 3.6 | 20 |
| Somatic kineties, number | 21 | 23 | 22 | 21.9 | 0.9 | 0.2 | 4.1 | 20 |
| Transverse ciliary rows, number | 10 | 11 | 11 | 10.9 | 0.4 | 0.1 | 3.4 | 20 |
| Macronucleus length | 21 | 26 | 23 | 23.1 | 1.4 | 0.3 | 6.1 | 20 |
| Macronucleus width | 17 | 23 | 21 | 20.5 | 1.7 | 0.4 | 8.3 | 20 |
TABLE 2. Morphometric comparison of Pinacocoleps spp.
| Species | Body shape | Body length, Number of Number of in vivo (μm) main plate teeth ciliary rows | Number of spines | References |
|---|---|---|---|---|
| P. tesselatus | Long oval-shaped | 75–85 AMP: 3, 21–23 PMP: 3 | PS: 3, AS: 2 | Present work |
| P. tesselatus | Barrel-shaped | 60–70 AMP: 3, 22–25 PMP: 3 | PS: 3, AS: 2 | Kahl (1930) |
| P. tesselatus * | Ovate to cylindrical | 46–49** AMP: 5, ca. 18 PMP: 3 | PS: 3, AS:? | Borror (1972) |
| P. i n c u r v u s | Curved cylindrical | 65–80 AMP: 7, 10–11 PMP: 7 | PS: 3, AS: 5 | Kahl (1930) |
| P. similis | Ovoid to barrel-shaped | 55–65 AMP: 5, 15 PMP: 4 | PS:5–7, AS:? | Chen et al. (2010) |
| P. heteracanthus | Long oval-shaped | 74–86 AMP: 9–11, ca. 22*** PMP: 7–9 | PS: 6***, AS: 3*** | Noland (1937) |
| P. arenarius | Rectangular | 70–75 AMP: 3, ca. 25*** PMP: 3 | PS: 10 AS: 1 | Borror (1952) |
| P. p u l c h e r | Long oval | 80–100 AMP: 10, 20–22 PMP: 10 | PS: 7, AS: 6 | Kahl (1930) |
| P. spiralis | Barrel-shaped | ca. 50 AMP: 7–8, ca. 23 PMP: 5–6 | PS: 3*** AS: 1*** | Noland (1937) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |