
Diplodinium cameli f. bispinatum, Kubesy, A. A. & Dehority, Burk A., 2002
|
publication ID |
https://doi.org/ 10.5281/zenodo.155871 |
|
DOI |
https://doi.org/10.5281/zenodo.5586600 |
|
persistent identifier |
https://treatment.plazi.org/id/996087B0-6D58-9946-7F64-50A4220CF0E6 |
|
treatment provided by |
Plazi |
|
scientific name |
Diplodinium cameli f. bispinatum |
| status |
f. n. |
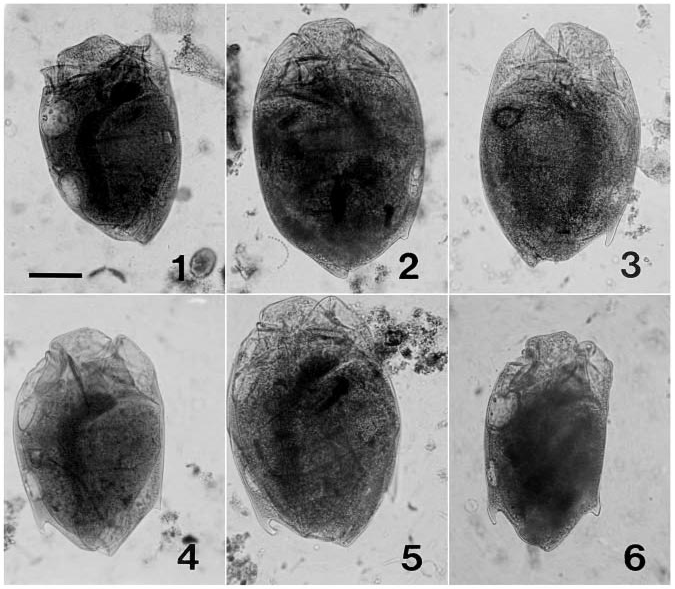
Diplodinium cameli f. bispinatum f. n. ( Figs. 46 View FIGURES 16 )
With all the characteristics of the species. Single spines arise from both the dorsal and ventral sides approximately fivesixths of the cell length towards the posterior end. The dorsal spine is well developed in most cells, averaging around 12 m, while the ventral spine ranges from about 5 to 12 m. The ventral spine tends to be slightly more pointed than the dorsal spine. This form only constituted 5.3% of Diplodinium cameli cells in the three animals in which it occurred. Dimensions for this form are presented in Table 3 View TABLE 3 .
Although there were some differences in size for the different forms, they were not significant except for the higher L/W ratio of D. cameli f. bispinatum (P <0.05).
The environmental or nutritional pressures which might lead to the development of spines in Diplodinium cameli are not known. Coleman, Laurie and Baily (1977) observed that in vitro cultures of Entodinium bursa required the presence of Entodinium caudatum , which they engulfed as a food supply. Addition of the nonspinated forms of Entodinium caudatum resulted in the development of spined cells. Their E. caudatum cultures had been previously grown for 17 years in vitro as the spineless form. Although development of spines probably requires additional energy compared to the nonspinated form, they found that ingestion of the spined form was very limited compared to the spineless form. They concluded that spination was actually a defense mechanism.
Because of its body size and the relatively small size of the spines, it would seem unlikely that Diplodinium cameli has developed the spines as a defense against predation. However, in ruminants the specific predation of large entodiniomorphs such as Eudiplodinium maggii by Polyplastron multivesiculatum has been well documented ( Eadie 1962, 1967). Other than this, most observations suggest that predation among the protozoa is accidental and very limited ( Lubinsky 1957). The absence of Polyplastron in the camels would seem to rule out the development of spination as a means to inhibit predation.
Van Hoven (1975), studying rumen protozoa in the tsessebe (antelope) from South Africa, reported the presence of spines in the species Diplodinium costatum . Later, Dehority (1985) observed spined forms of D. costatum in rumen contents from muskoxen in the Canadian arctic. Although Poljansky and Strelkow (1938) demonstrated that clone cultures in vivo of Entodinium caudatum were environmentally plastic and could be affected by diet it seems unlikely that this would explain the occurrence of spined forms in D. costatum . Diets would be quite different in these widely separated geographic locations. More recently, spined forms of Diplodinium rangiferi were observed in Australian red deer and in Japanese cattle which were inoculated with spineless forms of this species from sika deer ( Dehority 1997; Imai et al. 2002).
The spines observed in D. costatum and D. rangiferi cells are quite similar to those found in the different forms of D. anisacanthum ( Dogiel 1927) . That is, they arise at the caudal end of the cell. In contrast, the spines in D. cameli arise approximately onesixth of the distance toward the anterior end of the cell, from the dorsal and ventral surfaces.
The present study also revealed a wide variation in size, shape, and ciliary zones of Hsiungia triciliata and Polymorphella bovis , as well as several different forms of Entodinium ovumrajae . Further studies are required for possible redescription or establishment of new forms for these species.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |