
Eucorydia miyakoensis Yanagisawa, Sakamaki, and Shimano, 2021
|
publication ID |
https://doi.org/ 10.12782/specdiv.26.145 |
|
publication LSID |
lsid:zoobank.org:pub:E6F9409F-E720-437E-AA08-3D5E9542A49F |
|
persistent identifier |
https://treatment.plazi.org/id/2B08CF65-0790-4F3C-875C-4BDD1D1E34C6 |
|
taxon LSID |
lsid:zoobank.org:act:2B08CF65-0790-4F3C-875C-4BDD1D1E34C6 |
|
treatment provided by |
Felipe |
|
scientific name |
Eucorydia miyakoensis Yanagisawa, Sakamaki, and Shimano |
| status |
sp. nov. |
Eucorydia miyakoensis Yanagisawa, Sakamaki, and Shimano , sp. nov.
[New Japanese name: Benieri-rurigokiburi] ( Figs 1A–P View Fig , 2 View Fig )
Material examined. Holotype: male ( NMNS, NSMT-I- Dct-542), Gusukube , Miyako-jima Island, Okinawa, Japan, 12 November 2019, S. Yanagisawa leg. (eM-001) . Paratypes: 2 males and 5 females ( NMNS, NSMT-I-Dct-543–549), Gusukube , Miyako-jima Island, Okinawa, Japan, 12 November 2019, S . Yanagisawa leg . (eM-002–eM-008).
Differential diagnosis. This new species resembles E. guilinensis Qiu, Che, and Wang, 2017 , E. dasytoides , E. yasumatsui , E. tokaraensis , E. donanensis , and E. pilosa Qiu, Che, and Wang, 2017 . However, it can be easily distinguished by an uninterrupted transverse orange band in the middle of the tegmina, L7 of male genitalia round the basal half, R2 round, and a pair of orange pubescent blotches at the base of the tegmina in both males and females ( Fig. 1A–D, F–H, J, K View Fig ). Eucorydia miyakoensis sp. nov. can be distinguished from E. guilinensis and E. pilosa through characteristics of their genitalia. The L7 of male genitalia of E. guilinensi s is strongly protruded in the anterior and curved toward left posterior ( Qiu et al. 2017), whereas that of E. miyakoensis sp. nov. is round in the basal half ( Fig. 1F, H View Fig ). In addition, E. guilinensis resembles E. miyakoensis sp. nov. in that it has pubescence at the base of the tegmina; however, the pubescence of E. guilinensis is yellowish white, whereas that of E. miyakoensis sp. nov. is orange ( Fig. 1J, K View Fig ). Furthermore, the overall length of the male E. guilinensis is more than 13 mm (13.8–14.2 mm; Qiu et al. 2017), whereas that of the male E. miyakoensis sp. nov. is less than 13 mm (overall length of 12.5–13.0 mm; Fig. 1A–D View Fig ). The R2 of male genitalia of E. pilosa is elongated and slightly rhomboid ( Qiu et al. 2017), whereas that of E. miyakoensis sp. nov. is round ( Fig. 1G View Fig ). Eucorydia dasytoides and E. pilosa are distinguishable from the new species in that their males have an overall length> 18 mm ( Qiu et al. 2017), whereas overall length of the male E. miyakoensis sp. nov. is <13 mm. Both the size and shape of genitalia are similar to those of the three Japanese species, namely E. yasumatsui , E. tokaraensis , and E. donanensis , but it can be distinguished from them by orange pubescence at the base of the tegmina.
Description. Male (n=3): Body length 11.5 mm; overall length 12.5–13.0 mm; pronotum length 3.4–3.5 mm, width 5.6–5.9 mm; tegmen length 10.0– 10.8 mm ( Fig. 1A, B View Fig ).
Head shiny, black. Antenna black, consisting of 36–38 segments with 6–7 whitish subapical segments. Numbers of whitish segments sometimes different between the left and right sides. Pronotum metallic blue to metallic bluish green. Tegmina metallic blue to metallic bluish green, similar to pronotum color with a pair of orange pubescent blotches at the base near scutellum. Distal half with distinct, uninterrupted orange band. Hindwings hyaline, pale brown, becoming darker toward the apex with an orange blotch at middle of costa. Sc single; RA+RP with 5–8 branches; M single, but with one cell and some crossveins in middle part; CuA with 8 branches; CuP single; and AA+AP with 11–14 branches ( Fig. 1L View Fig ). Legs shiny, black.
Dorsum dark purple with yellowish area occupied from caudal half of 2nd segment to 5th segment; 6th segment of tergite dark purple in middle and yellow on lateral sides ( Fig. 1M View Fig ). Ventrum black, with yellowish area occupied from 2nd to 6th segments. Supra-anal plate black, widely bilobed; cercus consisting of 8 segments ( Fig. 1E View Fig ); subgenital plate black, rounded, with styli.
Genitalia ( Fig. 1F–I View Fig ): Left phallomere: L3 slender and curved, gradually narrowing toward the apex with a distinct hook ( Fig. 1I View Fig ); L7 round in the basal half, spatulate in the distal half, with beak-like apex ( Fig. 1H View Fig ). Right phallomere: R2 slightly elongated, round; basal left with a lobated protrusion ( Fig. 1G View Fig ).
Female (n=5): Body length 13.0– 13.7 mm; pronotum length 3.5–3.9 mm, width 6.1–6.5mm; tegmen length 8.7– 9.0 mm ( Fig. 1C, D View Fig ). Head, pronotum, and tegmen similar to male in color ( Fig. 1K View Fig ). Dorsum brownish black with yellow area occupied from 2nd to 5th segments and lateral sides of 6th segment. Supra-anal plate semicircular in shape and brownish black. Ventral side of abdomen brownish black with yellow area occupied from 1st through 6th segments; subgenital plate brownish black.
Ootheca (n=4): Length 3.0– 3.1 mm, width 5.0–6.0 mm. Light brown, trapezoidal pouch, with fine serration on one side of the margin with five longitudinal ridges on each side ( Fig. 1P View Fig ).
Larva (last instar): Male (n=1) body length 11.5 mm, mesonotum width 7.0 mm ( Fig. 1N View Fig ). Female (n=1) body length 12.1 mm, mesonotum width 7.5 mm. Body brown, with many fine bristles ( Fig. 1O View Fig ).
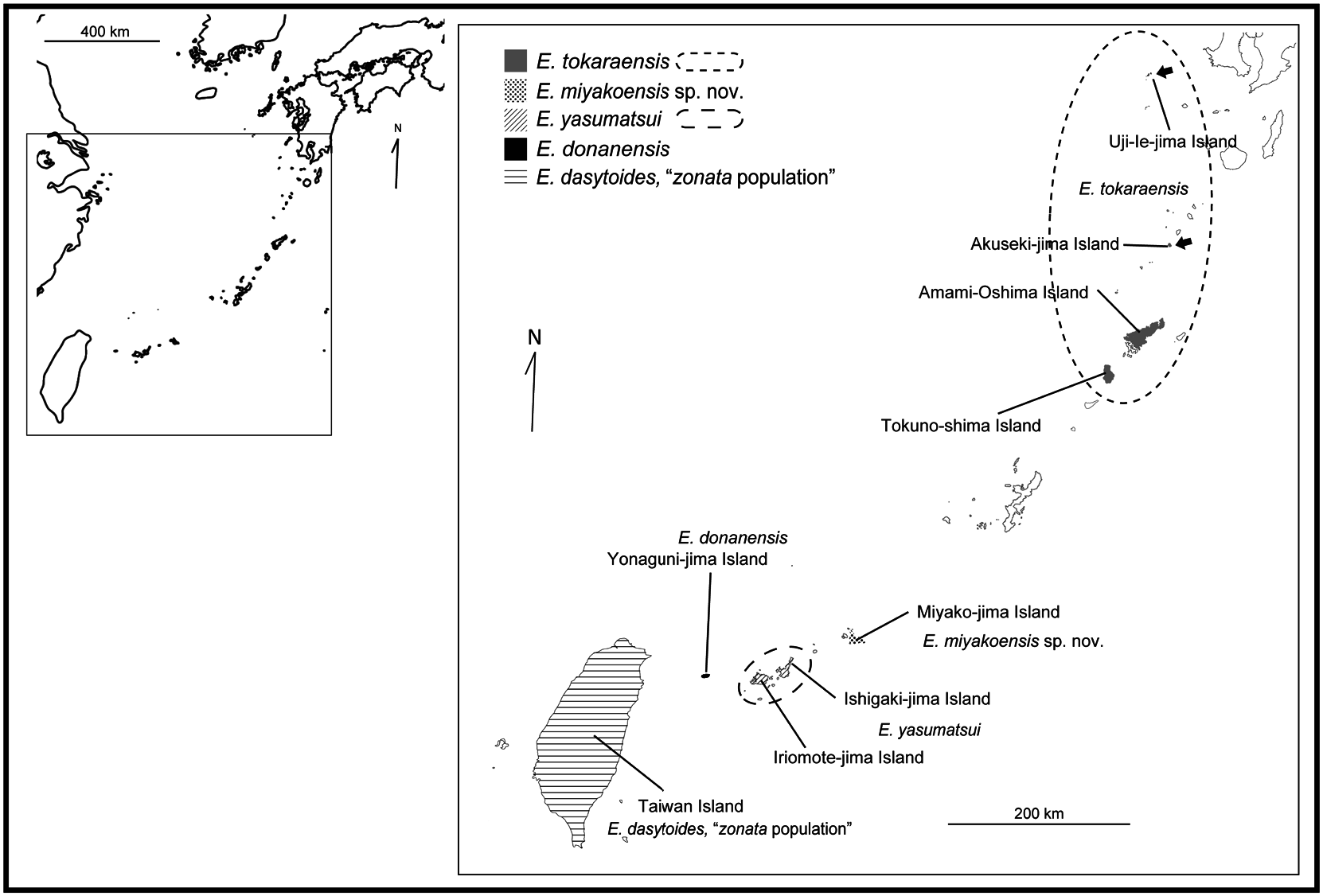
Distribution. Miyako-jima Island, Southwest Japan ( Fig. 2 View Fig ).
Etymology. The new species was named after the sampling site Miyako-jima Island.
Sequences. LC565385 View Materials – LC565406 View Materials , including two nuclear markers (28S and H3) and four mitochondrial markers (12S, 16S, COI, COII) from four specimens (eM-009–eM-012).
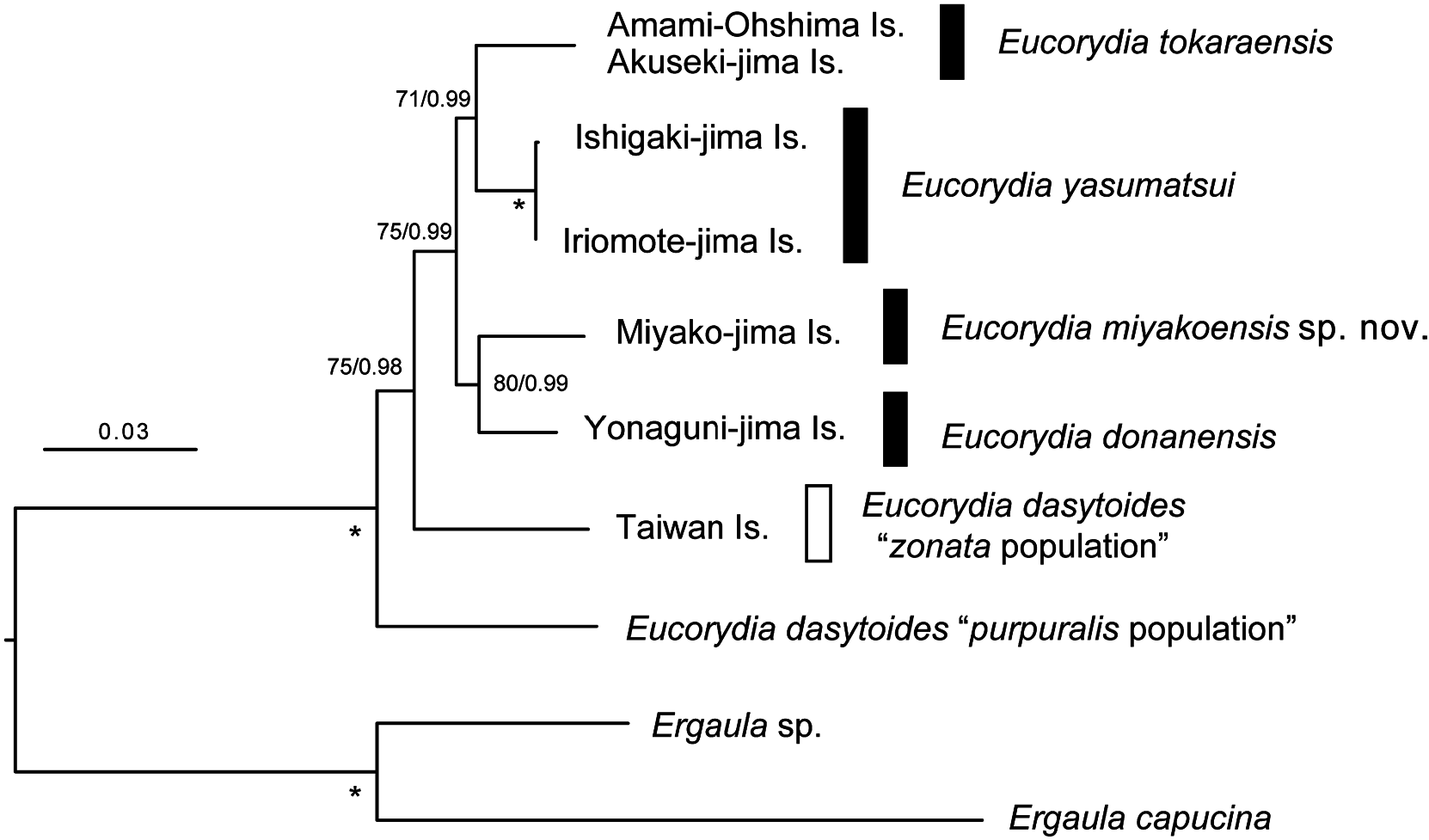
Phylogeny and genetic distance. Table 3 lists the sequences used for molecular phylogenetic reconstruction, including five molecular markers obtained from four individuals of E. miyakoensis sp. nov. ML tree inferred from the five concatenated markers is shown in Fig. 3 View Fig with nodal support values. Trees resulting from ML and BI had identical topologies; therefore, only ML tree is presented. The Japanese Eucorydia populations formed a sister clade with the E. dasytoides population of Taiwan Island. The Japanese populations were also monophyletic and divided into four lineages. New species from Miyako-jima Island formed a clade with E. donanensis from Yonaguni-jima Island.
Table 4 lists pairwise genetic distances of COI sequences in Japanese Eucorydia species and related species. Although only a small number of individuals were determined to have the COI sequence, Japanese Eucorydia species are genetically homologous on each island, and almost no intraspecific variation was detected for the COI gene. Only E. yasumatsui , distributed in Iriomote-jima and Ishigaki-jima Islands, has three haplotypes in COI sequences. Differences between these haplotypes include one or two substitutions (K2P 0.2%–0.3%). The new species were separated by genetic distances ranging from 3.4% to 6.7% (K2P) ( Table 3). Similarly, genetic distances between the Japanese Eucorydia species and the zonata population of E. dasytoides were relatively large (K2P 8.7%–10.3%). estimates are shown above the diagonal.
Remarks. This species was found only on Miyako-jima Island ( Fig. 2 View Fig ). The presence of this new species was also suggested in the remarks section which discussed the Eucorydia sp. (= E. tokaraensis ) in Asahi et al. (2016), but it was not described as a valid species since there were an insufficient number of specimens at that time. Obscureness of the orange band on the tegmina varied among individuals. The oothecae of this species are similar to the ootheca of E. yasumatsui described by Fujita and Machida (2014).

| NMNS |
National Museum of Natural Science |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |