
Eupolymnia chlorobranchiata, Nogueira, João Miguel Matos, Hutchings, Pat & Carrerette, Orlemir, 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4019.1.18 |
|
publication LSID |
lsid:zoobank.org:pub:906BB67C-F137-4CDA-A26B-77A025725800 |
|
DOI |
https://doi.org/10.5281/zenodo.5313625 |
|
persistent identifier |
https://treatment.plazi.org/id/039887C6-FFD2-9E31-FF31-4408FC5A0F51 |
|
treatment provided by |
Plazi |
|
scientific name |
Eupolymnia chlorobranchiata |
| status |
sp. nov. |
Eupolymnia chlorobranchiata View in CoL n. sp.
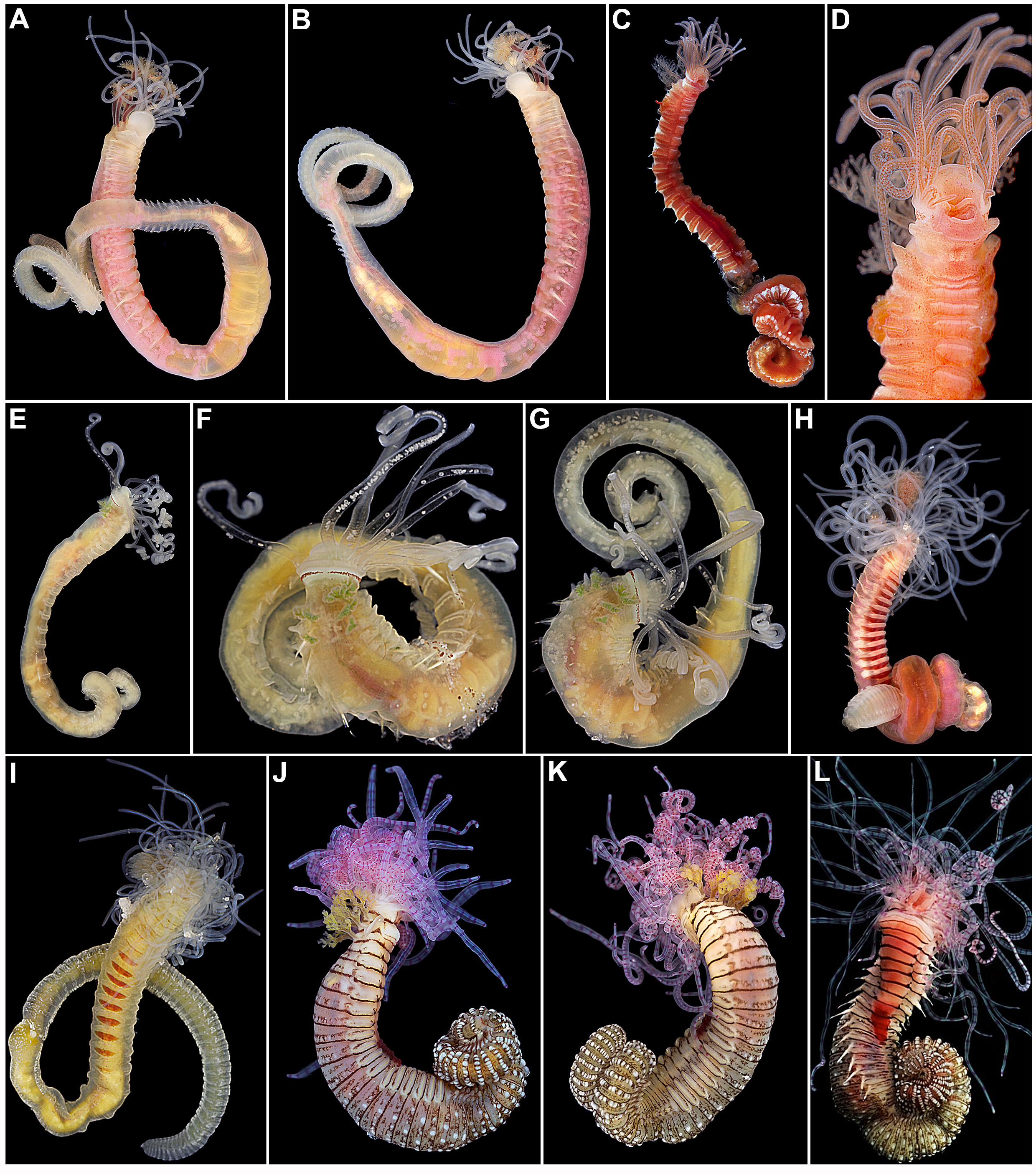
( Figs 2 View FIGURE 2 E–G, 36–38)
Eupolymnia koorangia View in CoL .— Hutchings & Glasby 1988: 13 (in part).
Type material. Holotype: AM W.47716, CReefs, LI–10–018, MI QLD 2193, Mermaid Cove (buoy), 14°38'46"S, 145°27'13"E, complete specimen, in 2 pieces, anterior end 6 mm long, 1 mm wide, posterior part 8 mm long, 1 mm wide. Paratypes: AM W.47801, CReefs, LI–10–018, MI QLD 2193, same locality as holotype; AM W.47717, CReefs, LI–10–062, MacGillivray Reef, deep reef slope, 14°39'25"S, 145°28'22"E, 2 posteriorly incomplete specimens 12 mm long, 2 mm wide; 6 mm long, 2 mm wide; AM W.47718, CReefs, LI–10–42, MI QLD 2204, North Point, 14°38'40"S, 145°27'17"E, complete specimen, 73 segments, ~ 16 mm long, 1.3 mm wide; AM W.45454, MI QLD 2446, mounted on 2 SEM pins; AM W.45455, MI QLD 2446 (photographed); AM W.47722, CReefs, LI–10–072, MI QLD 2209; AM W.47719, CReefs, LI–10–039, MI QLD 2203, incomplete specimen, more or less in good state of preservation, with 35 segments; AM W.47720 (2), CReefs, LI–10–047, MI QLD 2205, incomplete, with 23 segments, other incomplete, with 30 segments; AM W.47723, CReefs, LI–10–127, MI QLD 2232, incomplete specimen, with 30 segments, 9 mm long, 1.7 mm wide.
Other material examined. AM W.47760, CReefs, LI–10–035, Martin Reef, back reef, 14°45'22"S, 145°21'46"E, Aug 2010; AM W.44268, MI QLD 2340; AM W.45456, MI QLD 2446 (2); AM W.44525, MI QLD 2387; AM W.44515, MI QLD 2380; AM W.44969, MI QLD 2437 (4); AM W.44517, MI QLD 2381; AM W.44948, MI QLD 2413; AM W.44623, MI QLD 2413; AM W. 200728 – 200739, all from Lizard Island.
Comparative material examined. Holotype of Eupolymnia koorangia Hutchings & Glasby, 1988 , AM W.14028. Paratypes of Eupolymnia koorangia , AM W.14027, AM W. 200726.
Description. In life, colourless body and buccal tentacles, except for conspicuous row of prostomial eyespots and green branchiae ( Fig. 2 View FIGURE 2 E–G). Transverse prostomium attached to dorsal surface of upper lip; basal part with continuous row of eyespots without mid-dorsal gap, but with a single eyespot wide line instead mid-dorsally, with eyespots widely separated from each other ( Figs 2 View FIGURE 2 E–G; 36C, F–G); distal part shelf-like ( Figs 36 View FIGURE 36 A–C, E–G; 37A–J). Peristomium forming lips; hood-like upper lip, almost circular, about as wide as long; lower lip small, swollen, restricted to oral area ( Figs 36 View FIGURE 36 A–C, E–G; 37A–J). Segment 1 conspicuous all around, dorsally short, almost inconspicuous, ventrally developed, forming ventral lobe below lower lip; segment 2 with one pair of triangular, relatively wide ventro-lateral lobes, dorsal margins aligned with ventral edges of neuropodia; segments 3 and 4 with progressively shorter, triangular to semi-circular lobes, those of segment 3 almost same length as those of segment 2; both dorsal margins inserted very slightly progressively more laterally, almost aligned to each other and to line of notopodia ( Figs 36 View FIGURE 36 A–C, E–G; 37A–C, E–I). Paired dorso-lateral arborescent branchiae present on segments 2–4, first pair slightly dorsal to following pairs, which are longitudinally aligned; all dorsal to line of notopodia, with short branchial filaments branching dichotomously in few levels from short basal stem; first pair longer, about half body width of segment 2, other pairs about half size of first pair, slightly progressively shorter ( Figs 2 View FIGURE 2 E–G; 36A–C, E–G; 37A–J). Anterior segments with glandular, rectangular, smooth to slightly corrugated anteriorly mid-ventral shields, on segments 2–16, last shield distinctly shorter; mid-ventral groove extending posteriorly from segment 17 ( Figs 36 View FIGURE 36 A–B, E; 37A–C, E–I). Short and conical notopodia beginning from segment 4, extending for 17 segments, until segment 20, those of segments 4–9 originating progressively more laterally, then laterally aligned, notopodia of first 2 pairs, on segments 4–5, slightly shorter ( Figs 36 View FIGURE 36 A–G; 37A–K). Narrowly-winged notochaetae in both rows throughout, those of posterior row with wings on distal half ( Figs 36 View FIGURE 36 H–I; 38A–C). Neuropodia present from segment 5, as low ridges until termination of notopodia, on segment 20, as rectangular pinnules thereafter ( Figs 36 View FIGURE 36 A–B, D–E, G; 37A–C, E–H, K). Neurochaetae throughout as short-handled avicular uncini, arranged in completely intercalated double rows on segments 11–20; uncini throughout with elongate base, dorsal button at mid-length of base and crest with 2 transverse rows of secondary teeth, distal row with irregularly sized teeth; crest with more teeth per row after termination of notopodia ( Figs 36 View FIGURE 36 J–L; 37L–M; 38D–H). Nephridial papillae on segment 3, between bases of lobes and branchiae, genital papillae on segments 6–8, as round swellings between parapodial lobes, posteriorly ( Figs 36 View FIGURE 36 G; 37A–C, E–H, K). Pygidium papillate. Tube unknown.
Remarks. Capa & Hutchings (2006) provided a comparative table for all the species of this genus, which makes it clear that for many species data are inadequate available to really define them.
Three species, E. dubia ( Caullery, 1944) , E. intoshi ( Caullery, 1944) , E. robustus ( Caullery, 1944) have been described from Indonesia, E. koorangia Hutchings & Glasby, 1988 was described from SA and E. congruens ( Marenzeller, 1884) was from Japan. Eupolymnia chlorobranchiata n. sp. can be distinguished from E. congruens by the shape of the lateral lobes on segment 3, which in the latter species have a prolonged margin, absent in the former. Eupolymnia dubia and E. robustus have double rows of uncini on segments 10–20 whereas in E. chlorobranchiata n. sp. uncini are in double rows on segments 11–20. The other two species E. intoshi and E. koorangia differ from the E. chlorobranchiata n. sp. by the shape and arrangement of lateral lobes, which are arranged obliquely, and also their branchiae are not vertically aligned, while in E. chlorobranchiata n. sp. the branchiae of segments 3 and 4 are aligned longitudinally, as are the dorsal margins of lobes. For this reason we describe E. chlorobranchiata n. sp. as a new species, but a major revision of this genus is needed, with a revised description of the type species of the genus, E. nesidensis (Delle Chiaje, 1828) . Clearly, the shape and position of dorsal margins insertion of lobes are important in this genus, but descriptions of most species do not mention them. As apparently Delle Chiaje never deposited type material, the designation of a neotype for the type species should be a priority.
Type locality. Lagoon, 14°41'14"S, 145°27'18"E, Lizard Island, GBR, Australia.
Distribution. Widely distributed around the Lizard Island region.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Terebelliformia |
|
Family |
|
|
Genus |
Eupolymnia chlorobranchiata
| Nogueira, João Miguel Matos, Hutchings, Pat & Carrerette, Orlemir 2015 |
Eupolymnia koorangia
| Hutchings 1988: 13 |