
Hesiospina legendrei, Bonifácio & Lelièvre & Omnes, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4441.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:1E00CE34-E889-4514-A546-D17F6FFF86D2 |
|
DOI |
https://doi.org/10.5281/zenodo.5968103 |
|
persistent identifier |
https://treatment.plazi.org/id/DC771CF6-CD76-4A76-9CE9-6BB5F3A258AB |
|
taxon LSID |
lsid:zoobank.org:act:DC771CF6-CD76-4A76-9CE9-6BB5F3A258AB |
|
treatment provided by |
Plazi |
|
scientific name |
Hesiospina legendrei |
| status |
sp. nov. |
Hesiospina legendrei View in CoL sp. nov.
( Figs. 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 )
Type material: NE Pacific Ocean , Endeavour Segment (Juan de Fuca Ridge), Grotto site (47°56.958’N, 129°5.899’W), 2196 m depth, collected by ROV Jason 6–7 September 2015, Dive J 0831: Holotype GoogleMaps , MNHN-IA- TYPE 1845 , J0831-S1 incomplete specimen, length 13.27 mm, width 1.72 mm, 37 segments, accession GenBank MH349482 View Materials for 16S and MH349485 View Materials for COI ; Paratype 1, MNHN-IA-TYPE 1846, J0831-S3 incomplete specimen, length 11.51 mm, width 1.73 mm, 33 segments, accession GenBank MH349481 View Materials for 16S and MH349484 View Materials for COI ; Paratype 2, MNHN-IA-TYPE 1847, J0831-S1 complete specimen, length 11.17 mm, width 1.05 mm, 43 segments, accession GenBank MH349480 View Materials for 16S and MH349483 View Materials for COI ; Paratype 3, MNHN-IA-TYPE 1848, J0831-S3 complete specimen, length 13.94 mm, width 1.20 mm, 50 segments; Paratype 4, MNHN-IA-TYPE 1849, J0831-S3 complete specimen, length 12.18 mm, width 1.35 mm, 42 segments.
Additional material: NE Pacific Ocean, Endeavour Segment (Juan de Fuca Ridge), Grotto site (47°56.958’N, 129°5.899’W), 2196 m depth, collected by ROV Jason 6–7 September 2015, Dive J0831: Additional material 1 MNHN-IA-PNT 82, J0831-S3 incomplete specimen, length 12.99 mm, width 1.39 mm, 30 segments; Additional material 2 MNHN-IA-PNT 83, J0831-S3 incomplete specimen, length 11.33 mm, width 1.20 mm, 38 segments; Additional material 3 MNHN-IA-PNT 84, J0831-S3 incomplete specimen, length 11.46 mm, width 1.03 mm, 40 segments; Additional material 4 MNHN-IA-PNT 85, J0831-S2 incomplete specimen, length 13.65 mm, width 1.19 mm, 38 segments; Additional material 5 MNHN-IA-PNT 86, J0831-S2 incomplete specimen, length 12.28 mm, width 1.06 mm, 34 segments; Additional material 6 MNHN-IA-PNT 87, J0831-S2 incomplete specimen used to SEM. NE Pacific Ocean, Endeavour Segment (Juan de Fuca Ridge), Grotto site (47°56.958’N, 129°5.899’W), 2196 m depth, collected by ROV ROPOS 19 May 2014: Additional material 7 MNHN-IA-PNT 88, ONC 2014 R1689 incomplete specimen, length 11.36 mm, width 1.30 mm, 29 segments; Additional material 8 MNHN-IA- PNT 89, ONC 2014 R1689 complete specimen, length 19.55 mm, width 1.21 mm, 47 segments
Description (based on holotype and paratypes): Moderate-sized species, up to 19.55 mm long, up to 50 segments; dorsoventrally cylindrical anteriorly and posteriorly slightly flattened; live specimen not observed; ethanol preserved pale yellow to white; from segment 4–5, ventral-posterior part of neuropodia slightly inflated, with slight to dark brownish pigmentation.
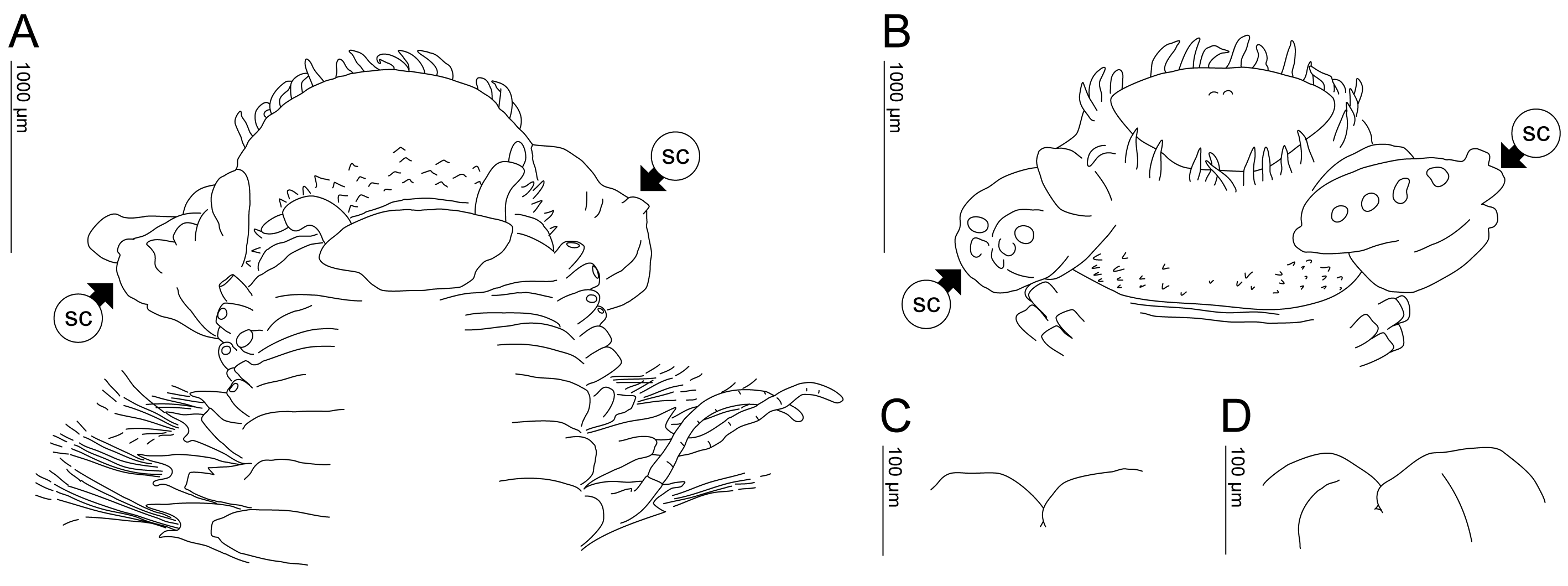
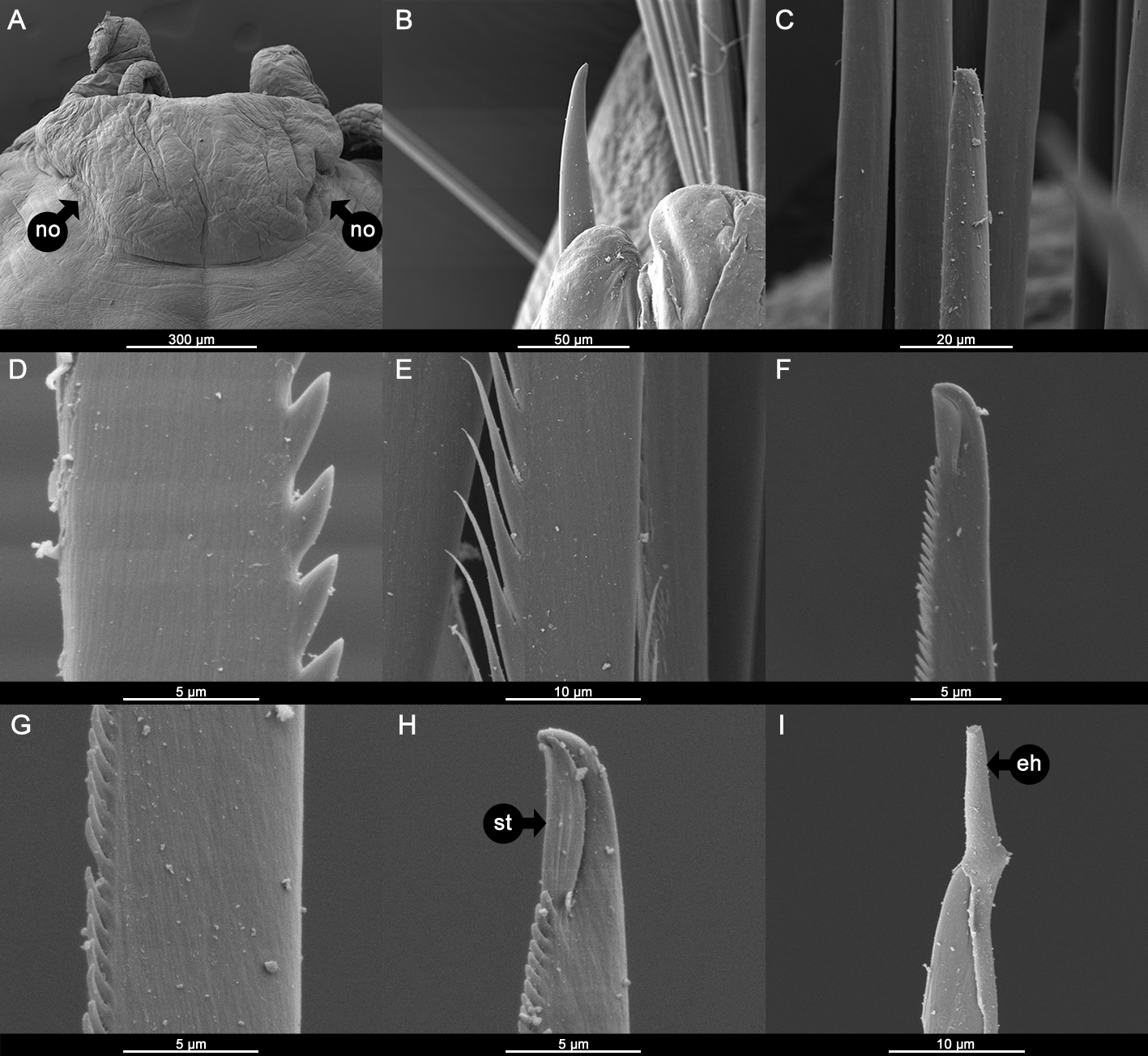
Prostomium wider than long, trapezoid, anteriorly wide, slightly notched posteriorly ( Fig. 2A View FIGURE 2 , 4A View FIGURE 4 ). Two antennae, smooth, thin, slightly tapering, long (as long as palpophores and palpostyle) with rounded tips, inserted on anterior margin close to palpophores; without median antenna. Palpophores cylindrical, slightly annulated, long (shorter than prostomium length) with palpostyle ovoid, thinner, shorter (about 1/3 of palphophores; Fig. 2A View FIGURE 2 ). Eyes absent. Nuchal organs as a patch of pores located laterally to prostomium ( Fig. 4A View FIGURE 4 ). Everted proboscis large, thick, bulbous (as long as the first 6 segments when everted; Fig. 2A View FIGURE 2 ); basally encircled with numerous, small, conical, pointed papillae (increasing in size distally), ranged in about 21–27 poorly defined vertical rows with 2–5 papillae by row; medially with a pair of large, sac-like structures, inflated, distally with 7–13 digitiform extensions, inserted ventro-laterally; distally with 20–27 elongate, thin, similar-sized papillae; not ciliated ( Fig. 2A, B View FIGURE 2 ); two pairs of small, delicate jaws, both pairs basally jointed, dorsal pair anteriorly rounded, ventral pair anteriorly slightly pointed ( Fig. 2C, D View FIGURE 2 ).
Dorsal cirri articulated, in segments 1–4 as tentacular cirri, very long (reaching 9 subsequent segments) with well-developed, cylindrical cirrophores; in subsequent segments, style shorter, long (reaching 2–5 subsequent segments) with smaller cirrophores.
Ventral cirri in segments 1–4 slightly articulated, long (reaching 4 subsequent segments) with well-developed, cylindrical cirrophores; in subsequent segments, style not articulated, slightly thinner, short (reaching distal neuropodia margin) with indistinct cirrophores, inserted subdistally ( Fig. 3A View FIGURE 3 ).
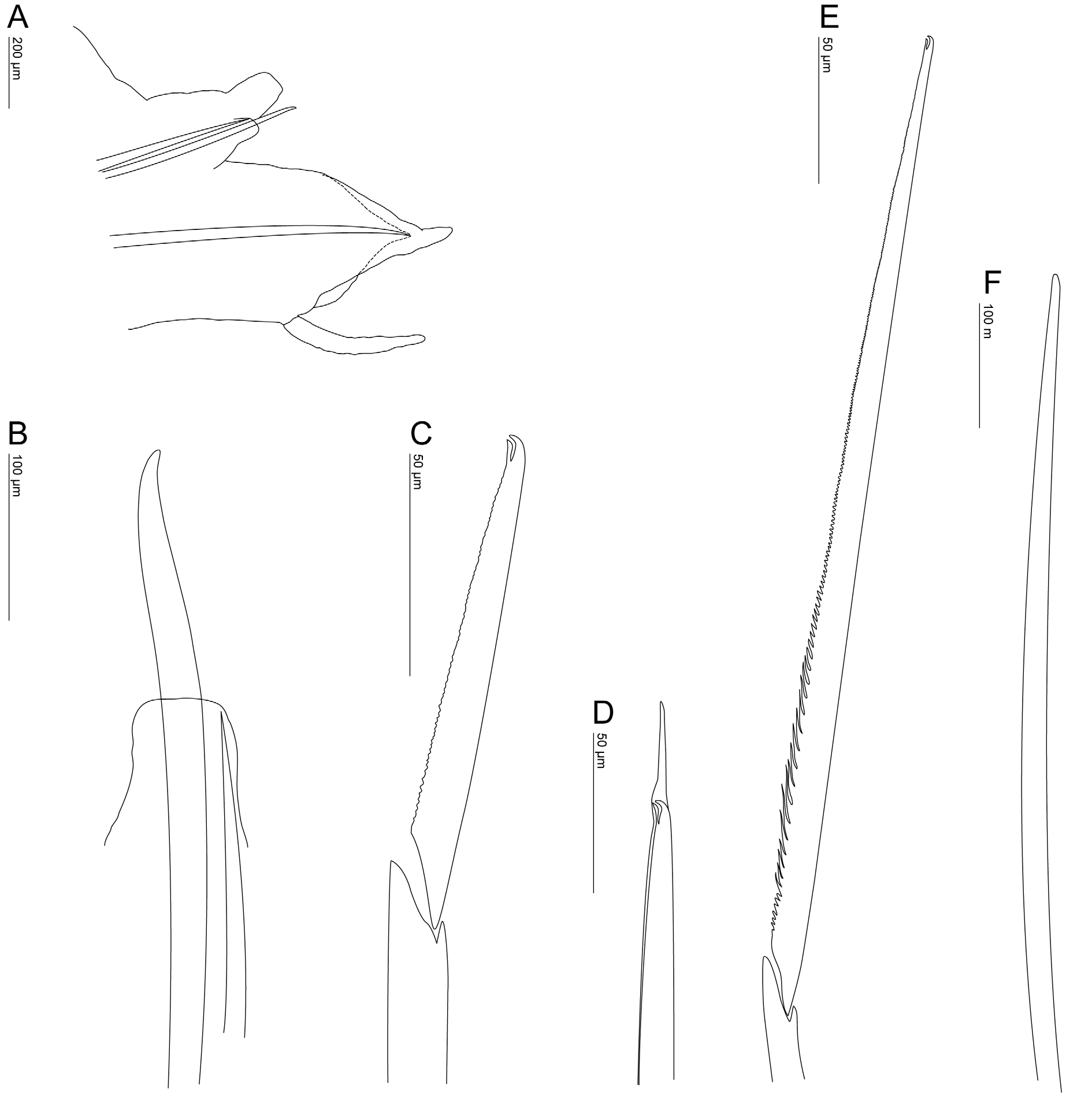
Parapodia subbiramous ( Fig. 3A View FIGURE 3 ). Notopodia reduced to small lobe; in segments 1–5 with 5–6 internal, slender aciculae; from segment 6 (chaetiger 2) to last segments with single internal, stouter acicula; from segments 7–8 (chaetigers 3–4) to last segments with single stout protruding hook (plate-like striae), very gently curved distally, with blunt to slightly pointed tips, emerging from small lobe located anteriorly to dorsal cirrophore ( Fig. 3A–B View FIGURE 3 , 4B View FIGURE 4 ).
Neuropodia well-developed, subtriangular, from segment 5 (chaetiger 1) to last segments; pre-chaetal lobe developed, triangular with pointed, prominent tips; post-chaetal not so developed, triangular, shorter, prominent ( Fig. 3A View FIGURE 3 ). Neurochaeta starting from segment 5 (chaetiger 1) to last segments of 2 types: (a) single stout, simple, long, slightly curved chaeta, with blunt tips, located closer to the neuroacicula ( Fig. 3F View FIGURE 3 , 4C View FIGURE 4 ); and (b) numerous (about 30–36 observed), slender, composite heterogomph falcigers with faintly serrated ( Fig. 3C, E View FIGURE 3 , 4G View FIGURE 4 ), tapering blades, with prolonged subdistal tooth and unidentate tips ( Fig. 3C, E View FIGURE 3 , 4F, H View FIGURE 4 ); medial blades at least 4 times longer than the inferiormost chaetae on fascicle, some with distinct, bigger or prolonged teeth series near base of blade ( Fig. 3E View FIGURE 3 , 4D–E View FIGURE 4 ); few (1–2 observed) inferiormost neurochaeta with elongated hood ( Fig. 3C–D View FIGURE 3 , 4I View FIGURE 4 ). All chaetae shafts, aciculae and hooks internally camerated (not drawn).
Pygidium as simple ring, bulbous, termino-dorsally located; last 2 segments very reduced.
Morphological variation: Few morphological variations were observed, for instance protruding hook appearing from segment 7 or 8 (chaetigers 3–4), occurred each case in half of analysed specimens. The number of distal papillae in the proboscis varied from 20 to 27 and seems to be linked to age-size (2 worms with everted proboscis). Few specimens in bad conditions presented the protruding hook accompanied externally by the internal acicula appearing to have “2 protruding hooks”. The holotype presented one segment with pre-chaetal lobe having slightly bifurcate tips in anterior region while the additional material 7 MNHN-IA-PNT 88 presented pre-chaetal lobe with bifurcate tips in few anterior segments. Furthermore, some specimens presented crenulated post-chaetal lobes without defined pattern, probably linked to an artefact of fixation.
Etymology: The new species is named in honour of Dr. Pierre Legendre (Université de Montréal) for his many contributions to marine ecology and deep-sea research.
Genetic data: DNA sequencing, for holotype and paratypes 1 and 2, was successful for 16S (404 to 421 bp length) and COI (504 to 687 bp length). Intraspecific variation for COI divergence was of 0.0% ( Table 2).
điagonal) anđ p-đistance (below điagonal) in MEGA7.
……continued on the next page ……continued on the next page
Distribution: Grotto edifice within Main Endeavour vent field (Juan de Fuca Ridge; 2196 m depth).
Ecological notes: Lelièvre et al. (2018) provided the first characterization of the biodiversity, community structure and trophic ecology in Grotto hydrothermal edifice (Main Endeavour, Juan de Fuca Ridge) which showed a site dominated by dense populations of tubeworms Ridgeia piscesae . Associated to this species, 35 other taxa can be found as well, mainly represented by gastropods and polychaetes, along the 6 sampled stations in this study. Among the 19 species of polychaetes observed associated to Ridgeia piscesae , Hesiospina legendrei sp. nov. was found in 3 out of 6 sampled stations with abundance varying between 9.8 and 14.3 ind.m -2. The same study found highest δ15N values and suggested the new species H. legendrei sp. nov. being among the top predators in the benthic food web.
Remarks: Together with Hesiospina aurantiaca and H. vestimentifera , H. legendrei sp. nov. is the third species described in the genus. Hesiospina aurantiaca —which lives mostly in shallow waters of the Atlantic, Caribbean, and western Pacific—can be easily differentiated from the hydrothermal vent taxa ( H. vestimentifera and H. legendrei sp. nov.) by the presence of a protruding hook distally curved, elongate palpostyles, eyes present, proboscis unarmed and absence of papillae in basal part of proboscis ( Pleijel 2004). Hesiospina vestimentifera and H. legendrei sp. nov. are both found in hydrothermal vent environments and share similarities such as: protruding hook slightly curved, ovoid palpostyle, eyes absent, proboscis with jaws and presence of papillae in basal part of proboscis. However, H. legendrei sp. nov. can be differentiated from H. vestimentifera , by having a sac-like structure ( Fig. 2A–B View FIGURE 2 ) present ventro-laterally in proboscis (absent in H. vestimentifera ). Furthermore, H. legendrei sp. nov. differs from H. vestimentifera , in having a greater number of distal papillae (about 20–27 observed) versus 13–14 ( Blake 1985) or 10–15 ( Pleijel 2004), notopodial hook from segment 7–8 (chaetigers 3–4) instead of segment 8–9 (chaetigers 4–5), and notopodia lobes with multiple slender aciculae in only the first 5 segments instead of the first 4 segments in H. vestimentifera . The elongate hood present in very few composite neurochaetae in H.
legendrei View in CoL sp. nov. seems to be a unique character among this genus but it should be investigated in the other species because it was rarely observed in our specimens. Moreover, genetic divergences support the new species with high Kimura 2-parameter distance (K2P) and p-distance for COI gene ( Table 2) varying respectively from 12.9 % and 11.8% between H. legendrei View in CoL sp. nov. and H. cf. vestimentifera View in CoL sp. 3 GR-2017 to 22.0% and 18.8% between H. legendrei View in CoL sp. nov. and H. vestimentifera View in CoL . It needs to be highlighted here two observations: (a) the type locality of H. vestimentifera View in CoL is Gálapagos Rift ( Blake 1985) but the only available DNA sequence was provided from one specimen sampled in Tui Malila, Lau Basin (about 1900 m depth; Pleijel et al. 2012); and (b) extreme values were observed between the sequence identified as H. cf. aurantiaca View in CoL MB-2010 and all others resulting in the exclusion of the COI sequence from this species of phylogenetic analysis.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Psamathinae |
|
Genus |
Hesiospina legendrei
| Bonifácio, Paulo, Lelièvre, Yann & Omnes, Emmanuelle 2018 |
legendrei
| Bonifácio & Lelièvre & Omnes 2018 |
H. legendrei
| Bonifácio & Lelièvre & Omnes 2018 |
H. legendrei
| Bonifácio & Lelièvre & Omnes 2018 |
H. cf. vestimentifera
| Blake 1985 |
H. vestimentifera
| Blake 1985 |
H. vestimentifera
| Blake 1985 |