
Hydroporus pervicinus Fall, 1923
|
publication ID |
https://doi.org/ 10.11646/zootaxa.1170.1.2 |
|
publication LSID |
lsid:zoobank.org:pub:87A2F77B-7BA8-47B1-B999-461C34D4A32E |
|
persistent identifier |
https://treatment.plazi.org/id/1C71EF58-1F59-847E-FEE3-FAEA39F0F918 |
|
treatment provided by |
Felipe |
|
scientific name |
Hydroporus pervicinus Fall, 1923 |
| status |
|
Hydroporus pervicinus Fall, 1923 View in CoL
Hydroporus pervicinus Fall, 1923: 84 View in CoL (orig. descr.); Anderson 1962: 63; Gordon & Post 1965: 17 (diagn.); Larson 1975: 311 (descr.); Larson et al. 2000: 399 (descr.); Nilsson 2001: 162 (cat.).
Hydroporus similaris Fall, 1923: 85 View in CoL (orig. descr.); Larson et al. 2000: 380 (descr.); Nilsson 2001: 166 (cat.); syn.n.
Hydroporus hirsutus Gordon, 1981: 117 View in CoL (orig. descr.); Larson et al. 2000: 401 (descr.); Nilsson 2001: 161 (cat.); syn.n.
Type material
Hydroporus pervicinus : Holotype : ♂ "Lake Tahoe \ Cal. \ Jul 17 21 97", " ♂ " [small label], "Type \ pervicinus." [partly hw Fall], "Type \ 23943" [red], " H.C. Fall \ collection", " Holotype \ Hydroporus pervicinus \ Fall 1923 " ( MCZ) .
Type locality: USA, California, Lake Tahoe .
Note: according to the original description there are paratypes from the same locality as the holotype and from British Columbia in SINM ( Sherman’s collection) as well as a single male from "Above Ouray, Colorado, Toll Road, 8000–9000 ft. " (Wickham’s collection). The paratypes have not been found in SINM and MCZ .
Hydroporus similaris : Holotype: ♂ "Corvallis \ Oreg", " ♂ " [small label], "Type. \ similaris" [partly hw Fall], "M.C.Z. \ Type \ 23954" [red], " H.C. Fall \ Collection", " Holotype \ Hydroporus \ similaris \ Fall 1923 " [hw] ( MCZ).
Type locality: USA, Oregon, Corvallis .
Note: according to the original description there are paratypes from the same locality as the holotype and one paratype from the Frazer [Fraser] Valley, British Columbia; all types were collected by Sherman. It is unknown where the paratypes are deposited. They have not found in SINM and MCZ .
Hydroporus hirsutus : Holotype: ♂ " Mt. Goethe \ Fresno Co. Calif. \ 9.VII.1952 \ Peter Raven" [partly hw], "12600 \ ft. elev." [hw], " Holotype \ Hydroporus hirsutus \ Robert Gordon", " California Academy \ of Sciences \ Type No. 9837" ( CAS).
Type locality: USA, California, Fresno County, Mount Goethe .
Note: according to the original description there are 15 paratypes from the same locality as the holotype which are deposited in CAS, SINM. The paratypes are with blue labels (personal communication with R. Gordon). The paratypes have not been found in SINM and I was not able to receive them from CAS .
Additional material
Canada: British Columbia: 3♂♂, 5♀♀ Blue Creek, Yalakom River , 6500’, 21.VIII.42, G.B. Leech ( CNC, CAS) ; 1♂, 1♀ Yalakom River, 21.VIII.42, Hugh B. Leech ( CNC) ; 1♀ E nderby, at brick factory, 19.VI.38, G.B. Leech ( CNC) ; 1♂ Mt. Apex , 5800’, 12.VIII.1933, A.N. Gartrell ( CNC) ; 1♂ Copper Mtn. , 10.VI.1929, G. Stace Smith ( CNC) ; 1♂ same locality only 17.IX.1929 ( CNC) ; 1♂ 21.VIII.1929 ( JBWM) ; 2♂ Coquihalla, Highway at Coldwater River , 16.IX.88, Carr ( CNC) ; 1 ex., Red Pass , 8.VIII.1932, G. Stace Smith ( CNC) ; 1♂, 5♀♀ Fraser Valley ( AMNH) ; 2♂♂, 1♀ Iskut, Highway 37, 11.VII.87, Carr ( CNC) . Alberta: 2♀♀ Manyberries , 21.VII.60, ( CNC) ; 1♂, 1♀ Cypress Hills, 20.V.61, Carr ( CGC, CNC) ; 1♂, 1♀ Cypress Hills , 23.VII.26, F.S. Carr ( JBWM) ; 1♂ Empress , 7.VI.1957, MacNay ( CNC) ; 1♂ Medicine Hat , 12.VI 1930, J.H. Pepper ( CNC) ; 1♂, 1♀ Medicine Hat , 31.VIII.24, F.S. Carr ( JBWM) ; 1♂ same locality only 31.V.24 ( JBWM) ; 1♂, 1♀ Aden, 20.VII.60, Carr ( CNC) ; 1 ♀ 2.5 mi SW Onefour , 22.V.1977, R.E. & M.L. Roughley ( JBWM) ; 1♀ Junction Highway 2 and Little Bow River , 9.IV.1971, D. & M. Larson ( JBWM) ; 3♂♂, 6♀♀ N Cressday, Twp. [township] 6, Rge. [range] 1, W 4 Mer. [meridian], 10.05.1980, Carr ( CHS) ; 2♂♂ Twp. 6, Rge. 1 W 4 Mer. [south Cypress Hills], 10.V.1980, Carr ( CNC) ; 1♂ Twp. 6, Rge. 7 W 4 Mer. [west Orion], 31.III.1972, Carr ( CNC) ; 2♀♀ " Twp. 13, Rge. 4 W 4 Mer. [east Medicine Hat], 21.V.1961, Carr ( CGC, CNC) ; 1♂ Twp. 18, Rge. 26 W 4 Mer. [near Brand], 23.IV.1972, Carr ( CNC) ; 1♂ Twp. 1, Rge. 12 W 4 Mer. [southeast Masinasin], 2.VII.1972 Carr ( CNC) ; 1♂ Twp. 28, Rge. 2 W 4 Mer. [near Sibbald], 20.VI.1985, Carr ( CNC) ; 1♂ Gleichen , 25.III.1956, Carr ( CNC) ; 1 ex., Twp. 6, Rge. 3 W 5 Mer. [west Beaver Mines], 6.VII.1961, Carr ( CNC) ; 1 ex., 4 mi N Lundbreck , 12.VII.1971, Larson ( JBWM) ; 2♂♂ Twp. 13, Rge. 4 W 5 Mer. [west Claresholm], 12.VII.1961, Carr ( CNC) ; 1♂ Calgary , 24.III.1956, Carr ( CNC) ; 1♂ Calgary , 3.V.1953, Carr ( CNC) ; 1♀ Calgary , 10.IV.1944, E.J. Kiteley ( CNC) ; 2♂♂ Jumpingpound Creek , 22.IV.1972, Carr ( CNC) ; 2♂♂, 3♀♀ 15 mi SE Calgary, 4.IV.71, D. & M. Larson ( CNC) ; 11♂♂, 7♀♀ near East Coulee, Twp. 28, Rge. 17 W 4 Mer. , 29.III.1986, Carr ( CHS) ; 1♂, 1♀ Piegen Indian Reserve , 23.VI.26, F.S. Carr ( JBWM) ; 2♂♂ Twp. 25, Rge. 3 W 5 Mer. , 2.V.1980, J. Carr ( CGC) ; 1♀ Wisdom [?], 21.VII.60, Carr ( CNC) . Saskatchewan: 2♂♂ Twp. 8, Rge. 27 W 3 Mer. [near Cypress Hill Provincial Park], 22.VIII.1986, Carr ( CNC) ; 4♂♂ Twp. 8, Rge. 28, W 3 Mer. [W Cypress Hill Provincial Park], 21.08.1986, Carr ( CHS) ; 1♂ Twp. 6, Rge. 2 W 3 Mer. [Twelve Mile Lake], 22.VII.1986, Carr ( CNC) ; 1♀ Twp. 6, Rge. 2 W 3 Mer. , 21.VI.1990, Carr ( CNC) ; 1♂ Twp. 8, Rge. 5 W 2 Mer. [near Kisbey], 1.VIII.1994, Carr ( CNC) ; 1♂ Lake Alma, 27.IV.87, Carr ( CNC) ; 1♂ Assiniboia , 10.VII.1973, Carr ( CNC) ; 1♂ Assiniboia , 7.VIII.1971, Carr ( CNC) ; 1♂ Regina , 14.V.1944, C.C. Shaw ( CAS) ; 1♀ Roche Percee , 4.VIII., J.B . Wallis ( CNC) . Manitoba: 14♂♂, 2♀♀ Highway 3, 2.8 km N junction with 256N, near Pierson , 19.IX.1985, R.E. Roughley ( JBWM, CHS) ; 1♂ Aweme , 30.VII.22, J.B . Wallis ( CNC); 2♂♂, 2♀♀ Aweme , 6.VII.20, J.B . Wallis ( CNC, JBWM) ; 1♀ Aweme , 30.VII.1922, N. Criddle ( CNC) ; 1♂, 1♀ near Reston , 28–29.VII.02, Shaverdo H. & Alperin M., 49º40’48"N 101º12’46"W ( CHS) GoogleMaps ; 1♀ Winnipeg , 24.V.24, J.B . Wallis ( JBWM) ; 1♀ same locality only 22.VI.24 ( JBWM) ; 1♀ Twp. 7, Rge. 10 E, 19.XII., J.B . Wallis ( JBWM) .
USA: Washington: 1♂, 1♀ Ritzville, 17.V.1921, M.C. Lane ( JBWM); 1♂, 8 exs., Highway 23, 3 km N Lamont, 12.VI.84, Carr ( CNC); 1♂ Reardan, 18.IV.83, Carr ( CNC). Montana: 1♂, 1♀ Highway 323, 45 mi N Alzada [Carter Co.], 23.VII.90, Carr ( CNC). Note: the species was previously reported from the state by Gordon (1969): Glacier Co. and Meagher Co. The records confirm that the species occurs in Montana. North Dakota: 3♂♂, 9♀♀ Stutsman Co., Cottonwood Lake, 2.IV.1980, B.A. Hanson & G.A. Swanson (NDSUe); 1♀ same locality only 18.IV.1980 (NDSUe); 1♀ 18.IV.1980 (NDSUe); 1♂, 1♀ 2.V.1979 (NDSUe). Note: the species was previously reported from the state by Gordon & Post (1965): Sioux Co. and by Gordon (1969): Sioux Co., Williams Co., and Towner Co. The records confirm that the species occurs in North Dakota. Oregon: 1♂, 1♀ "Corvallis [Benton Co.], 20.V.1938, Hugh B. Leech ( CAS); 1♂ Corvallis 22.IX. ( AMNH); 1♂ Corvallis, 15.III.1936, N.P. Larson ( AMNH); 1♂ Eugene [Lane Co.], 26.VI.41, B. Malkin ( CNC); 3♂♂, 12♀♀, 13 exs., Wasco Co., Mt. Hood National Forest, FSR 4310 at Clear Creek, 5.VI.1989, C.G. & D.A. Pollock ( JBWM, CHS); 6 exs., Wasco Co., Mt. Hood National Forest, FSR 4310 at Clear Creek Campground,. 5.VI.1989, D.A. Pollock ( JBWM); 3♂♂, 13♀♀ Steen [Steens] Mt. Road, South Limb [Harney Co.], 23.VI.84, Carr ( CNC); 7♂♂, 12♀♀ Highway 140 at Blue Creek [Lake Co., near Lakeview], 25.VI.84, Carr ( CNC, JBWM); 3♀♀ Seneca [Grant Co.], 18.VI.84, Carr ( CNC); 6♂♂, 3♀♀ Silver Lake [Lake Co.], 1.VII.84, Carr ( CNC, CHS); 3♂♂, 1♀ Highway 205, 20 mi N Fields Junction [Harney Co.], 20.VI.84, Carr ( CNC); 1♀ same locality only 21.VI.84 ( CNC); 1♀ 11 mi, 22.VI.84 ( CNC); 3♂♂, 1♀ Barton Lake Reservoir [Clackamas Co.?], 19.VI.84, Carr ( CNC); 7♂♂, 3♀ Silvies [Grant Co.], 18.VI.84, Carr ( CNC, CHS); 6♂♂ Bend, Swamp Wells, Deschutes National Forest, 3.VII.84, Carr ( CNC); 2♂♂, 6♀♀, 1 ex., Highway 205, Roaring Springs Ranch, 20.IV.84, Carr ( JBWM); 2♂♂ same locality only 22.VI.84 ( CNC); 1♂, 7♀♀ 10 mi E Blizzard Gap, Highway 140, 24.VI.84, Carr ( CNC); 1♂ Alkali Lake, 30.VI.84, Carr ( CNC); 1♂ Klamath Co., Poe Valley, 13.V.1966, Joe Schuh ( AMNH); 1♂ Barkley Spring, Upper Klamath Lake, 6.VI.1960, Joe Schuh" ( AMNH); 1♂ "Lake Co., 14 mi SW Plush, 6.VI.1958, Vertrees & Schuh ( AMNH). Idaho: 1♂ South Fork, Partridge Creek at Fish Creek Road, Targhee National Forest, 18.VI.86, Carr ( CNC); 1♂ Junction Roads 294 & 315, Targhee National Forest, 10.VI.86, Carr ( CNC); 4♂♂, 2♀♀ 27 km S Prairie, 31.V.86, Carr ( CNC, JBWM). Wyoming: 2♂♂ South Pass City [Fremont Co.], 7.VII.70, Carr ( JBWM). Note: the species was previously reported from the state by Gordon (1969): Sheridan Co. and Yellowstone National Park. The records confirm that the species occurs in Wyoming. California: 1♂ Lower Klamath Lake, Ore. [California], 30.V.55, Joe Schuh ( AMNH); 1♀ Siskiyou Co., Medicine Lake, 22.IX.1965, Joe Schuh ( AMNH); 1♂ Yuba Pass [Nevada Co.], 1.VII.91, Carr ( CNC); 1♂, 1♀ Tahoe [Placer Co.], VII. ( AMNH); 1♂ Siskiyou Co., 6 mi S Macdoel, 2.VII.1956, Joe Schuh" ( AMNH); 1♀ Yosemite National Park [Mariposa Co.], 17.VIII.79, Fery ( CHF). Note: see also " Type material". Nevada: 1♂ ½ mi E Spooners Summit, 15.VI.1957, P.C. & R.W. Coleman ( CAS); 3♂♂ White Pine Co., Snake Range, Wheeler Park, Stella Lake, 10800’, 29.VII.71, D.H. Kavanaugh ( CAS); 2♂♂ White Pine Co., Snake Range, Great Basin National Park, Stella Lake, 10.400’, 12.VIII.1994, M.A. Ivie ( MSUB); 3♀♀ Nevada ( CNC, AMNH). Utah: 1♀ Wasatch National Forest, 12.VIII.66, Carr ( CNC); 1♀ Manila, 8.VII.70, Carr" ( CNC); 1♀ Spirit Lake, Uinta Mountains, 30.VI.86, Carr ( CNC); 1♂, 1♀ Virgin River [Washington Co] ( AMNH); 1♀ Dixie National Forest, near Panquitch Lake, 9.VII.2004, Wewalka ( CGW). Colorado: 3♂♂, 2♀♀ Gove Pass, 11.VI.70, Carr ( CNC); 2♂♂ Mancos, 23.VII.70, Carr ( CNC); 1♂ Clear Creek Co., Berthoud Pass, 22.VII.69, O. Bazoska ( CNC); 1♀ Boulder Co., Rollins Pass, 18.VIII.1981, A.C. Ashworth, elevation 10800’ (NDSUg); 3♂♂, 2♀♀ Nederland, Science Lodge, 11500’, 4.VII.61, B.H. Poole ( CNC); 2♂♂ same locality only 27.VI.61 ( CNC); 1♂, 3 exs., Garfield Co., White River Mts., Trappers Lake, 9800’, 14–15.VIII. 73, D.H. Kavanaugh ( CAS). Note: see also " Type material". Arizona: 1♂, 1♀ Coconino Co., near Woods Canyon Lake, 4.VI.1983, J. Webb ( CGC); 2♂♂, 5 exs., KaibabLodge, 18.VII.66, Carr" ( CNC); 2♂♂, 4♀♀ Grand Canyon, North Rim, 8000–9100 ft, 19. VII.34, D. Rockefeller ( AMNH); 9♂♂, 7♀♀ Kaibab National Forest, N Grand Canion, 14.7.2004, Wewalka ( CGW); 16 exs., Mint Springs, Coconino National Forest, 26.IX.80, Carr ( CNC); 6 exs., Kehl’s Spring, Coconino National Forest, 27.IX.80, Carr ( CNC); 12 exs., Coconino Co., on Route 160, 2 to 3 mi E Mogollon Rim, 18.IX.68, P. Bartholomew ( CAS); 1♀ Coconino Co., head of Tonto Creek, Cn. [canyon] Mogollon Mesa, 2350 m, 16, 21.VIII.1977, G.E. & K.E. Ball ( JBWM); 1♂ Apache National Forest, 10 mi SW Eagar, 16.VII.76, 2600 m, M. Campbell ( CNC); 1♂ Gila Co., Sierra Ancha Mts., Aztec Peak, ca. 6.1 km SE junction 288 & Tonto National Forest Road 487, sfacing slope, 2220 m, 31.VIII.1977, G.E. & K.E. Ball ( JBWM); 2♂♂ Cospino ( AMNH).
Description
TL=3.56–4.64 mm, MW=1.80–2.32 mm, MW/TL–HL=0.52–0.57; habitus broadly oval to rather narrow and elongate ( Figs. 5–8 View FIGURES 1–13 ), somehow more convex than H. longiusculus , especially pronotum.
Elytra paler than head and pronotum or coloration of dorsal surface uniformly brownish black with reddish brown lateral margins of pronotum; head reddish brown to black, with paler vertex, ventrally yellowish brown to dark brown, usually genae distinctly darker than gula (gula reddish, genae dark brown) or slightly darker then gula (gula reddish brown, genae dark reddish brown), sometimes genae darker than gula only close to sutures; pronotum reddish brown to black, with broadly paler margins; elytra uniformly reddish brown to black, sometimes with yellowish spots at base ( Fig. 8 View FIGURES 1–13 ); pronotal and elytral epipleura pale reddish brown to brown; antennomeres not concolorous, antennomeres 1–2 yellowish red to reddish brown, not infuscate, antennomeres 3–4 usually infuscate, antennomeres 5–11 dark brown to black, paler at their base; palpi pale reddish brown (like gula) to brown, infuscate apically or not; legs yellowish brown to paler or darker reddish brown, tibia and tarsi darker; ventral side brownish black with reddish brown posterior margin of metacoxal processes and usually with pale reddish brown to dark brown lateral spots on abdominal sternum 3 or 3–4.
Head with punctation relatively coarse and dense (spaces between punctures 1–3 or 5 times size of punctures); disc of pronotum with punctation similar or denser than on head, with distinct, punctate impression at posterolateral angles; elytra with denser (spaces between punctures 1–2 times size of punctures), coarser, and more regular (not even) punctation than on disc of pronotum; epipleuron of elytron with large punctures, spaces between them 1–2 times size of punctures; metasternum, metacoxae, and abdominal sterna 1–2 with very large punctures, their medial part with very fine and sparse punctation, abdominal sterna 1–2 with punctation distinctly denser than on metasternum and metacoxae; abdominal sterna 3–6 with punctures much smaller and sparser than on abdominal sterna 1–2, abdominal sternum 6 with punctation rather variable among and within populations, from relatively sparse to rather dense; punctures with hairs; dorsal and ventral surfaces with evident microreticulation; most specimens with rather fine microreticulation, so that dorsal surface appearing shiny, some specimens with microreticulation stronger, thus appearing dull.
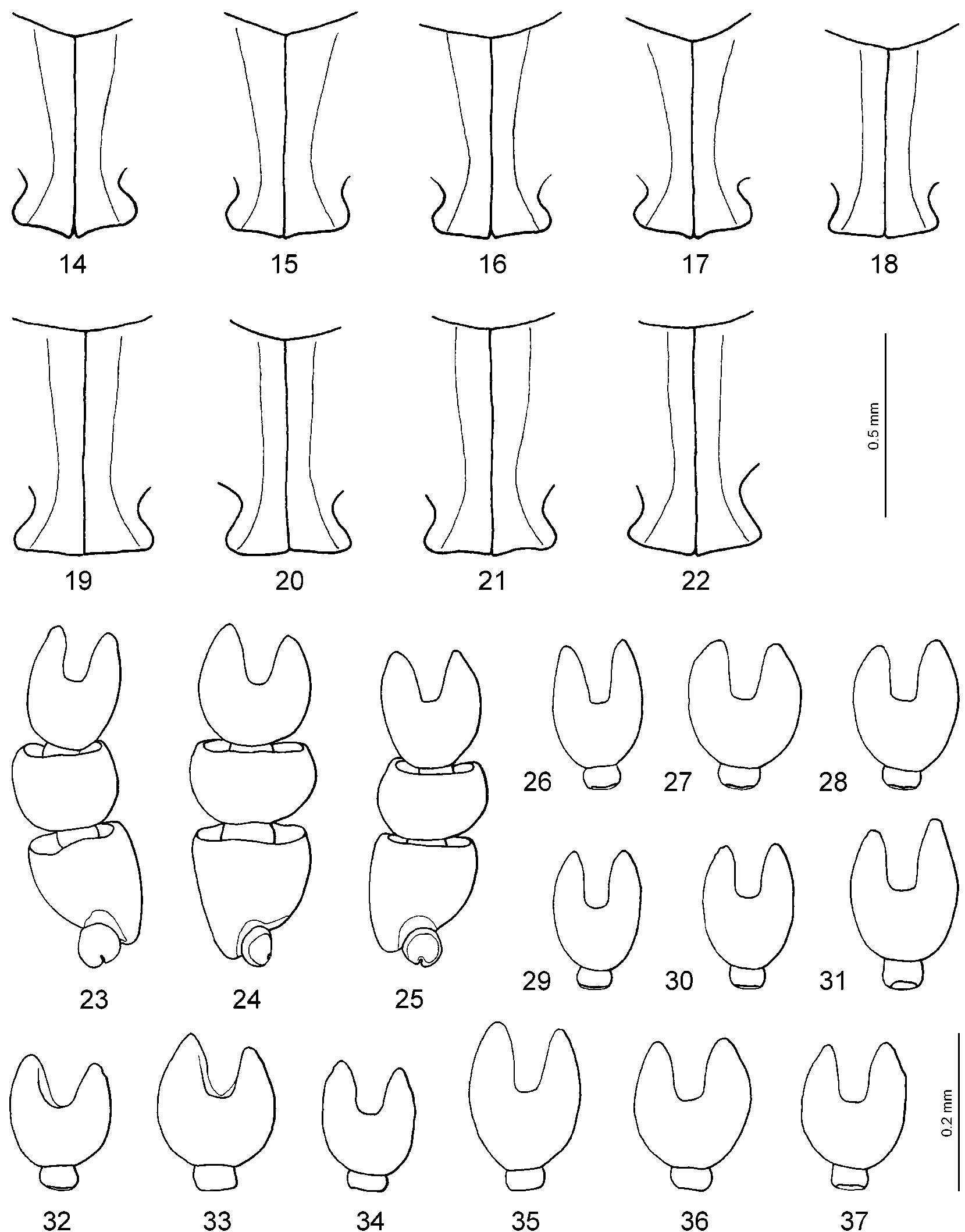
Pronotum with evident, relatively broad lateral bead; sides of pronotum rounded and convergent; epipleura slightly visible at anterior angles in lateral view; pronotum slightly sinuate at its base so that posterolateral angles slightly but distinctly produced. Lateral margin of elytron weakly to moderately ascending toward shoulder. Prosternum with declivity of prosternal process distinct and prominence evident. Metacoxal lines parallel or subparallel, in rare specimens slightly diverging; posterior margins of metacoxal processes conjointly truncate to slightly angulate, in some specimens more sinuate than angulate ( Figs. 19–22 View FIGURES 14–37 ).
Anterior surface of protibia with single puncture row which irregular basally—similar to row in H. longiusculus but in most specimens not so extreme ( Figs. 11–13 View FIGURES 1–13 ).
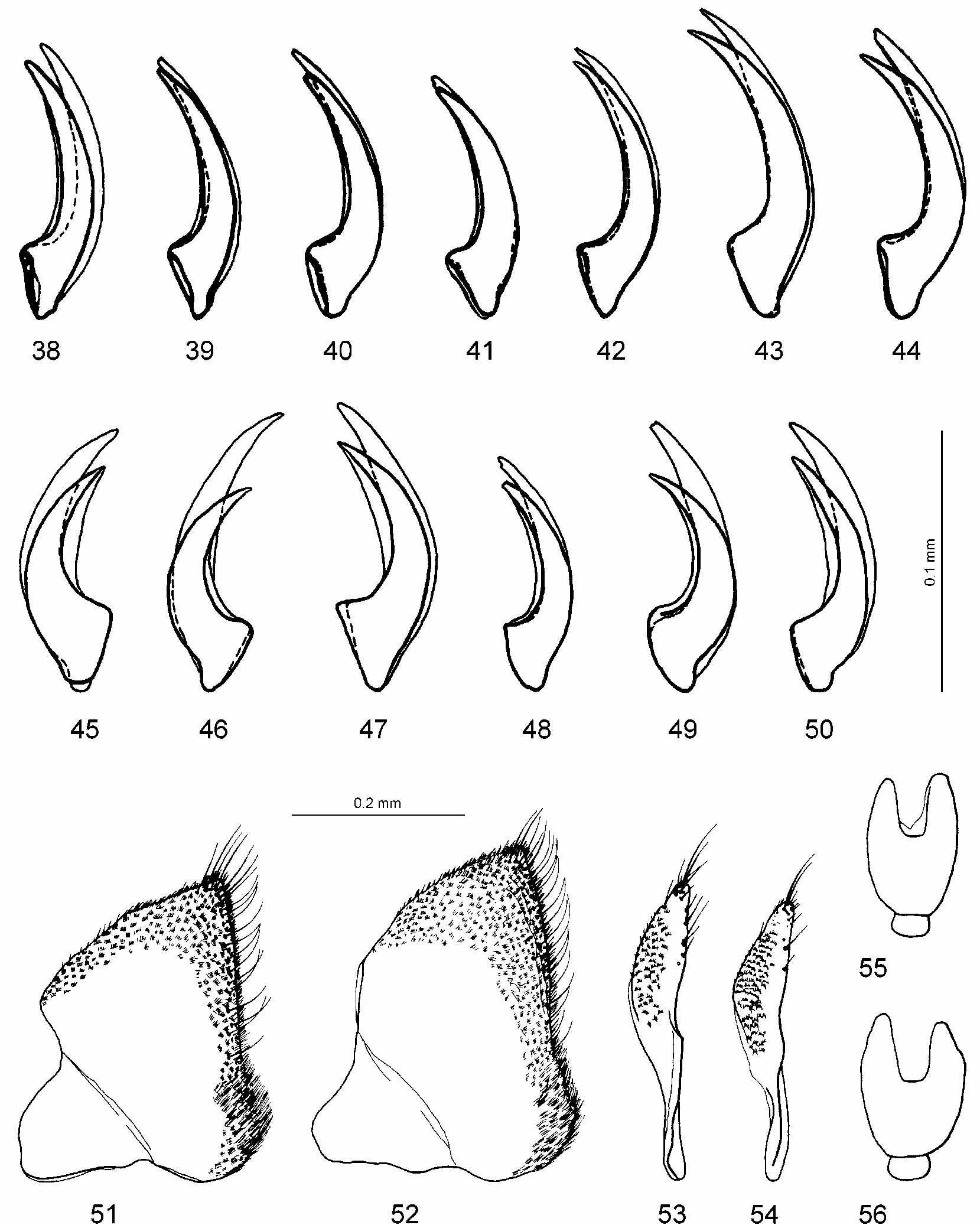
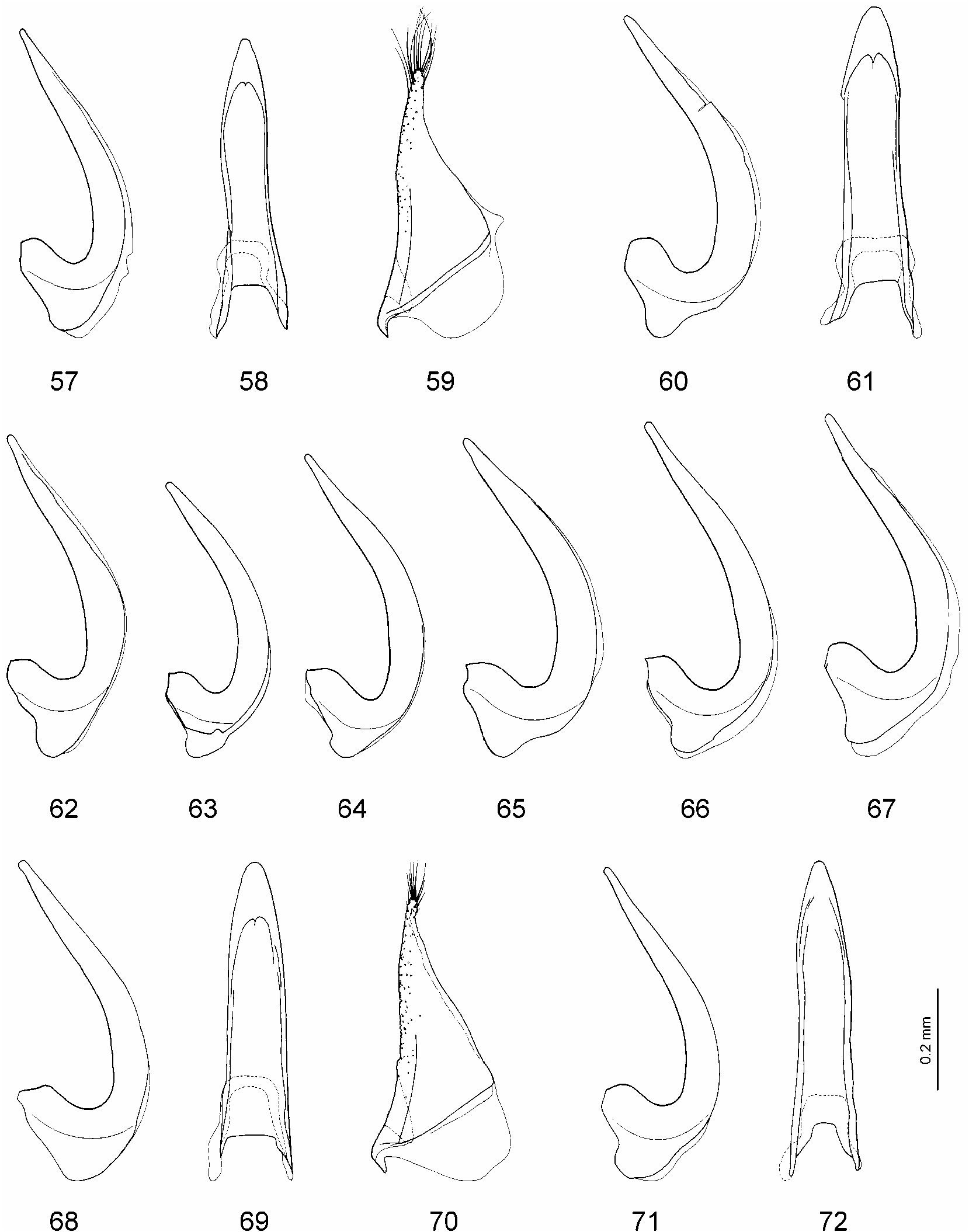
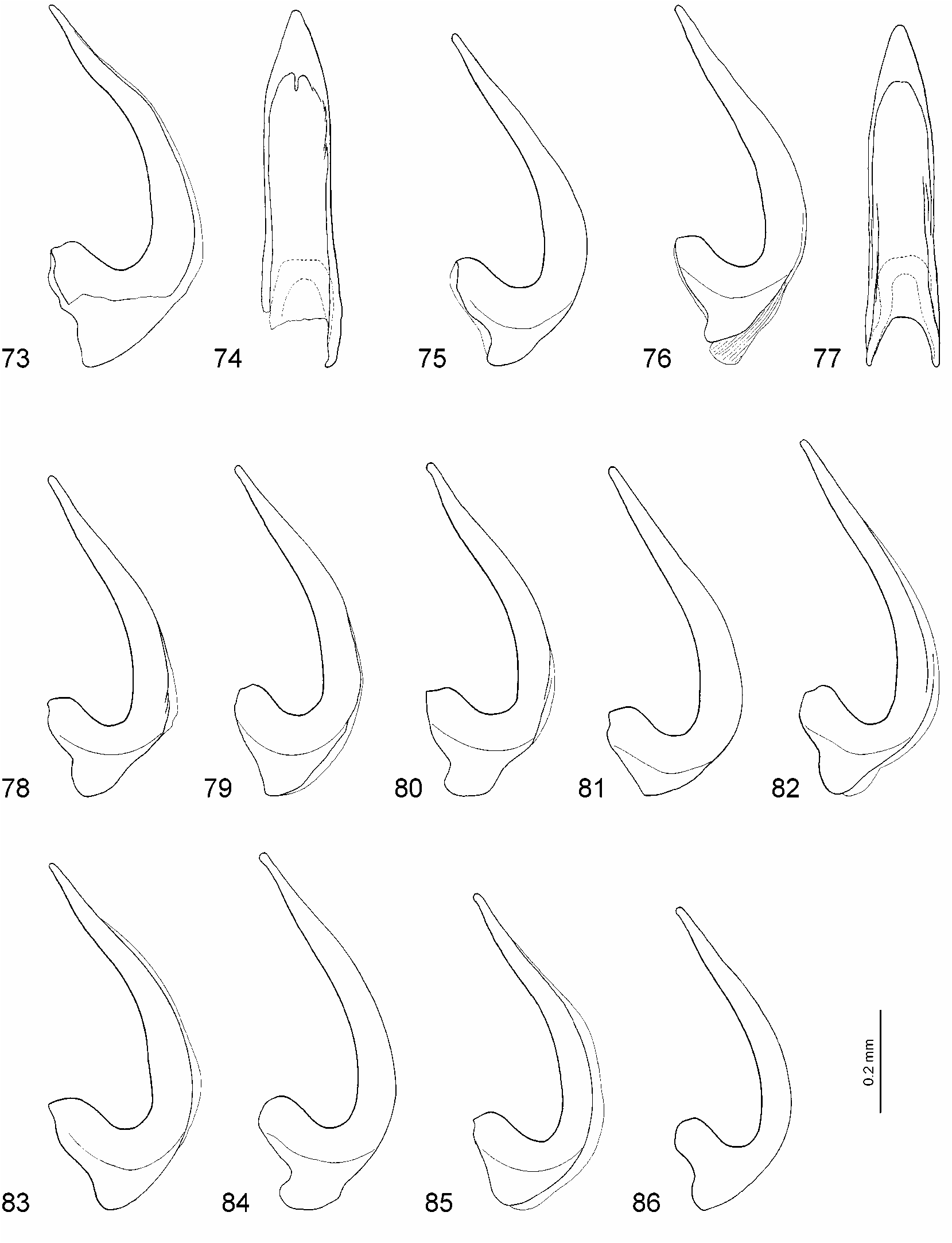
Male: Protarsomeres 1–3 dilated, rather small, variable in shape (oval or rounded) and size: Lprot3=0.14–0.19 mm, Lprot2=0.09–0.12 mm, protarsomere 3 the narrowest: Wprot3=0.13–0.15 mm, Wprot2=0.14–0.18 mm, Wprot1=0.14–0.16 mm, Wprot3/ Lprot3=0.74–1.00 ( Figs. 24, 25, 32–37 View FIGURES 14–37 ); protarsus with claws distinctly unequal, the anterior one evidently shorter (2/3) and thicker than the other, in some specimens less unequal in length and often in thickness ( Figs. 45–50 View FIGURES 38–56 ). Median lobe of aedeagus straight to curved in lateral view ( Figs. 68, 71 View FIGURES 57–72 , 73, 75, 76, 78–86 View FIGURES 73–86 ), in ventral view broadly pointed to pointed ( Figs. 69, 72 View FIGURES 57–72 , 74, 77 View FIGURES 73–86 ). Paramere as in Fig. 70 View FIGURES 57–72 .
Female: Protarsomere not modified, narrow, sometimes broader than in H. longiusculus ( Fig. 56 View FIGURES 38–56 ). Protarsal claws not modified, equal. Dorsal microreticulation same as in male or slightly stronger, sometimes females more shiny than males. Gonocoxosternum and gonocoxa as in Figs. 52, 54 View FIGURES 38–56 .
Variability
The species shows great variability in body shape, coloration, punctation, microreticulation, shape of posterior margin of the metacoxal processes, shape of male protarsomere 3, relative size of the protarsal claws, and shape of median lobe of the aedeagus.
The shape of the body varies from broadly oval to elongate as well as from more convex to flatter. Most of the studied specimens have oval body shape. Specimens from Oregon display a body shape from broadly oval to rather narrow and elongate. Single beetles from British Columbia have a slightly discontinuous body outline due to a pronotum with rounded and protruded sides. Beetles from one population (27 km S Prairie Id 31. V.89 lot 1 Carr) in Idaho have the body more convex and narrower or broader and flatter.
The coloration of the dorsal surface of the body varies from not uniform, with darker head and pronotum and paler elytra to uniformly black (or rarely reddish brown), with reddish lateral margin of pronotum. These two colorations are characteristic for specimens from all studied regions, except North Dakota where beetles are uniformly black, with lateral margin of the pronotum not distinctly paler. Some specimens from Washington, Oregon, Idaho, and Utah have yellowish spots at the base of the elytra similar to H. longiusculus ( Fig. 8 View FIGURES 1–13 ).
The punctation of the dorsal surface of the body varies from sparser (e.g. in specimens from Manitoba) to denser (e.g. in specimens from Washington and Oregon). Also the microreticulation is differently developed (independently male or female), even in specimens from the same population (Oregon) so that the dorsal surface of the body can be from shiny to rather dull. These two forms are characteristic for the specimens from all studied regions. For example in males from Saskatchewan two examples are shiny, two with microreticulation of elytra more strongly developed, and two rather dull, with strongly developed microreticulation on the elytra and pronotum. Specimens with more strongly developed microreticulation have been more often observed in southern populations.
The shape of the posterior margin of the metacoxal processes varies in specimens from all studied regions. For instance in beetles from Oregon it is truncate, in some specimens concave on both sides so that it s slightly sinuate to slightly angulate, and in some very slightly rounded ( Figs. 19–22 View FIGURES 14–37 ).
The male protarsomere 3 varies in size and shape. In most specimens from Manitoba, Saskatchewan, and Alberta it is medium sized and squarerounded ( Fig. 24 View FIGURES 14–37 ). In beetles from more southern populations it is more variable, from small to large and from squarerounded to elongate ( Figs. 25, 33–37 View FIGURES 14–37 ). In British Columbia and Oregon specimens with smaller protarsomere 3 are more frequently observed ( Figs. 32, 34 View FIGURES 14–37 ). In Oregon the protarsomere 3 is larger and more rounded or smaller and narrower in specimens from the same population ( Figs. 33, 34 View FIGURES 14–37 ). Also the shape of the protarsomere 3 is found often variable in beetles from Colorado ( Figs. 36, 37 View FIGURES 14–37 ).
Distinctly unequal male protarsal claws are characteristic for most studied beetles from different populations ( Figs. 45–47, 49 View FIGURES 38–56 ). However, beetles with less unequal claws (anterior one is less short) have been observed in the populations from Manitoba, Alberta, British Columbia, Wyoming, North Dakota, Idaho, Oregon, California ( Fig. 48 View FIGURES 38–56 ), and especially from Colorado, Arizona, and New Mexico. The specimens from Colorado have the claws distinctly unequal to almost equal in the same population ( Figs. 49, 50 View FIGURES 38–56 ).
The median lobe of the aedeagus varies from almost straight in lateral view to conspicuously curved, with the apex more or less pointed in ventral view. The straight shape of the medial lobe is characteristic for most studied specimens. A slightly curved shape of the medial lobe has been observed in specimens from Manitoba, Saskatchewan, British Columbia, Colorado, Utah, Arizona ( Figs. 75, 76, 78, 80, 85, 86 View FIGURES 73–86 ), and especially from Oregon. The most sinuate shape is characteristic for specimens from Corvallis (Oregon) ( Figs. 73, 83 View FIGURES 73–86 ). Also in Oregon specimens variability of the shape of the medial lobe from straight to distinctly sinuate is very well expressed ( Figs. 81–83 View FIGURES 73–86 ).
Remarks on the symonymy
Hydroporus hirsutus View in CoL was described by Gordon (1981), who wrote that the new species resembles H. pervicinus View in CoL and can be separated from it by the piceous last segment of the maxillary palpus, the much finer punctation of the metasternal plate, and nearly equal male protarsal claws. Larson et al. (2000) found that the holotype does not agree well with Gordon’s description (it is smaller and the median lobe of the aedeagus is not constricted medially, which was obviously the same problem as with H. utahensis View in CoL ). They wrote that H. hirsutus View in CoL "has no distinctive characters other than the conspicuous pale pubescence and somewhat obscure punctation of the metacoxal plane" as well as that the species is extremely close to H. pervicinus View in CoL with the median lobe and protarsomeres indistinguishable, only the anterior protarsal claw of the male is slightly modified. My comparison of the holotype of H. hirsutus View in CoL with numerous specimens of H. pervicinus View in CoL from different localities showed that it is conspecific with H. pervicinus View in CoL . The coloration of the last segment of the maxillary palpus is not a reliable character since it varies in H. pervicinus View in CoL from reddish brown with infuscate apex to uniformly reddish brown, brown, and black. Even the holotype of H. pervicinus View in CoL has brownish black last segments of the maxillary palpi. The punctation of the metasternum and metacoxae is a variable character, too. I examined specimens of H. pervicinus View in CoL from Manitoba and British Columbia that also had a rather fine and obscure punctation on these sclerites. The anterior protarsal claw of the male is really not so strongly modified as in "normal" H. pervicinus View in CoL , but it is distinctly shorter and more strongly curved than the posterior ( Fig. 48 View FIGURES 38–56 ). If the variability of this character in H. pervicinus View in CoL is taken into consideration, one cannot fully rely only on it for distinguishing the species. Some studied specimens of H. pervicinus View in CoL have the conspicuous pale pubescence like in H. hirsutus View in CoL . The body size and shape, coloration, and pubescence of the holotypes of H. hirsutus View in CoL and H. pervicinus View in CoL are very similar ( Figs. 6, 7 View FIGURES 1–13 ).
Hydroporus similaris View in CoL was described by Fall (1923) together with H. pervicinus View in CoL . He assumed that it resembled "most closely its near neighbor hirtellus View in CoL ". In my opinion, H. similaris View in CoL is conspecific with H. pervicinus View in CoL , though it was never placed close to this species by the previous authors ( Larson et al. 2000). My examination of the holotype showed that the species is similar to H. pervicinus View in CoL in the body shape, slightly but distinctly protruding posterior angels of the pronotum, slightly irregular anterior puncture row on the protibia ( Fig. 5 View FIGURES 1–13 ), and unequal male protarsal claws. The last character was mentioned in the original description: "the anterior claw in the male is barely or scarcely as long and slightly thicker than its fellow", as well as by Gordon (1981: 117) "…but similaris View in CoL males have the anterior protarsal claw distinctly shorter than the posterior". The only character that might indicate not being conspecific with H. pervicinus View in CoL is the shape of the median lobe of the aedeagus. It is slender and more curved in the apical part in lateral view ( Fig. 73 View FIGURES 73–86 ) and more pointed in ventral view ( Fig. 74 View FIGURES 73–86 ). However, in my opinion, it is within the variability range of this character. I have studied three males (one of them was identified by Leech as H. pervicinus View in CoL ) from the same locality (Corvallis, Oregon) as the holotype. One of them is teneral and the other two have a similar shape of the median lobe as in the holotype ( Fig. 83 View FIGURES 73–86 ). Some specimens from other localities in Oregon and from Manitoba, Saskatchewan, British Columbia, Colorado, and Arizona, also have a curved median lobe, though not so distinct as in the specimens from Corvallis ( Figs. 75, 76, 78, 80, 85, 86 View FIGURES 73–86 ). What is more, the median lobe of the holotype is a bit damaged laterally at the apex that probably makes the curvature slightly stronger. In ventral view the shape of the median lobe is also variable ( Figs. 69, 72 View FIGURES 57–72 , 74, 77 View FIGURES 73–86 ). In addition, eight specimens (five females and three males) from Fraser Valley, British Columbia were studied. These specimens bear old locality labels "Franz Val BC". I assume that the paratype of H. similaris View in CoL is also with such a label. Four females and two males were identified by Gordon as H. hirtellus View in CoL , one female —as H. despectus rusticus . The male without identification label surely belongs to H. pervicinus View in CoL . It has distinctly unequal protarsal claws and a slightly curved median lobe. All specimens have an irregular puncture row on the anterior surface of the protibia and the characteristic body shape of H. pervicinus View in CoL with slightly protruding posterior angels of the pronotum. Therefore, none of them belongs to H. despectus View in CoL . I consider that the females belong rather to H. pervicinus View in CoL than to H. hirtellus View in CoL since they are smaller, narrower, and slightly more convex than the two males of H. hirtellus View in CoL , of the same shape as the male of H. pervicinus View in CoL . Besides, the pronotum is darker than the elytra and the puncture row of the protibia is slightly irregular, whereas the two males of H. hirtellus View in CoL are uniformly coloured and with the puncture row strongly irregular.
As a result of the facts mentioned above I believe that H. similaris View in CoL should be considered a synonym of H. pervicinus View in CoL .
Ecology
According to Larson et al. (2000) the species occurs along margins of small, often temporary, grassland ponds—one of the few grassland species, which in Alberta shows a higher level of habitat association with Hygrotus species than with other species of Hydroporus . In Manitoba I collected this species in a grassland pond strongly overgrown with Typha and with much decaying vegetation. In Colorado H. pervicinus is recorded from a pond at tree line, at altitude ca. 3300 m. In Arizona the species was collected in a creek in the conifer forest, at altitude 2350 m. It is also known from streams, brooks, rivers, pools, and alkaline lakes.
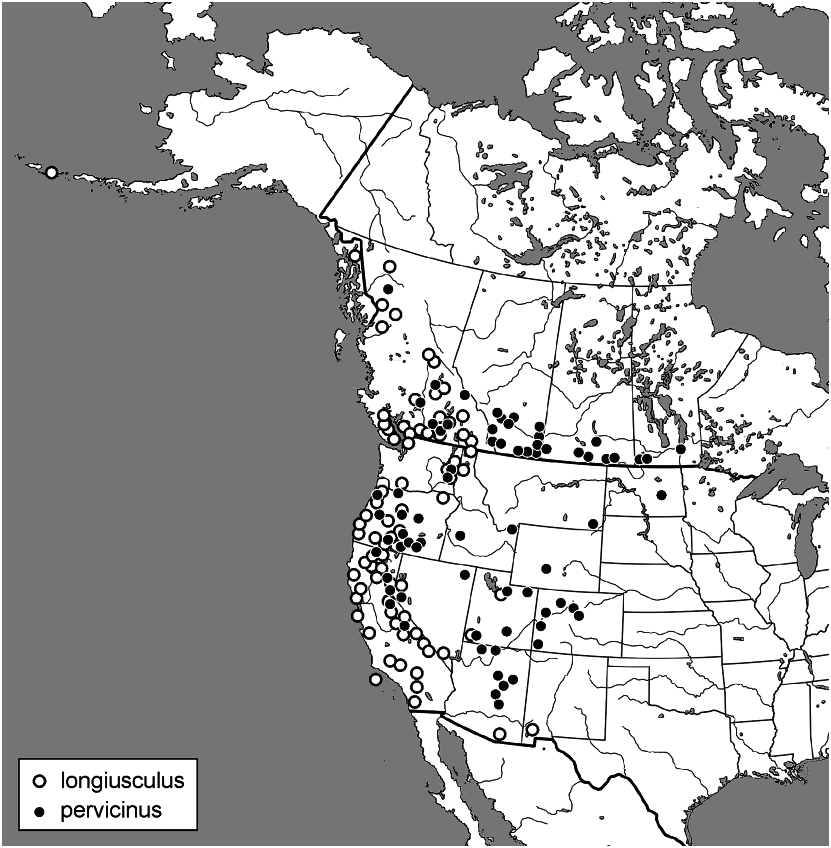
Distribution
CANADA: Manitoba, Saskatchewan, Alberta, British Columbia. USA: Washington, Montana, North Dakota, Minnesota: Bengal, Hibbing, St. Paul ( Wallis 1973) , Oregon, Idaho, Wyoming, California, Nevada, Utah, Colorado, and Arizona ( Fig. 87 View FIGURES 87 ).

| MCZ |
Museum of Comparative Zoology |
| CAS |
California Academy of Sciences |
| R |
Departamento de Geologia, Universidad de Chile |
| CNC |
Canadian National Collection of Insects, Arachnids, and Nematodes |
| JBWM |
J.B. Wallis Museum of Entomology |
| AMNH |
American Museum of Natural History |
| CGC |
Caenorhabditis Genetics Center |
| MSUB |
Montana State University |
| V |
Royal British Columbia Museum - Herbarium |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Hydroporus pervicinus Fall, 1923
| Shaverdo, Helena V. 2006 |
Hydroporus hirsutus
| Nilsson, A. N. 2001: 161 |
| Larson, D. J. & Alarie Y. & Roughley, R. E. 2000: 401 |
| Gordon, R. D. 1981: 117 |
Hydroporus pervicinus
| Nilsson, A. N. 2001: 162 |
| Larson, D. J. & Alarie Y. & Roughley, R. E. 2000: 399 |
| Larson, D. J. 1975: 311 |
| Gordon, R. D. & Post, R. L. 1965: 17 |
| Anderson, R. D. 1962: 63 |
| Fall, H. C. 1923: 84 |
Hydroporus similaris
| Nilsson, A. N. 2001: 166 |
| Larson, D. J. & Alarie Y. & Roughley, R. E. 2000: 380 |
| Fall, H. C. 1923: 85 |