
Iberorhyzobius rondensis ( Eizaguirre, 2004 ) Eizaguirre, 2004
|
publication ID |
https://doi.org/ 10.5281/zenodo.173892 |
|
DOI |
https://doi.org/10.5281/zenodo.6262080 |
|
persistent identifier |
https://treatment.plazi.org/id/03AA0541-A722-011A-0317-BECFFE002391 |
|
treatment provided by |
Plazi |
|
scientific name |
Iberorhyzobius rondensis ( Eizaguirre, 2004 ) |
| status |
comb. nov. |
Iberorhyzobius rondensis ( Eizaguirre, 2004) , comb. nov.
Type material (not studied by the authors): SPAIN: Holotype (S1) Puerto de Alijar (410 m, altitude), (Sierra de Palmitera, provincia de Málaga, Comunidad de Andalucia, 11.viii.1997); Paratypes (specimens S2S10), collection of Eizaguirre; all the specimens were collected in coniferous (species not mentioned by the author).
Additional material: PORTUGAL: Ferraria (Peninsula de Setubal): 5 adults obtained (iii–iv.2000) from larva collected in 20.ii.2000, on the trunk of Pinus pinaster trees infested with Matsucoccus feytaudi (1ɗ, PCAR; 4Ψ, EFN); Sintra ( Monserrate): 6 adults collected in 15.vi.2003 (1ɗ PCCC; 1ɗ, 4Ψ EFN); 1 adult obtained (iv–v.2004) from larva, collected idem (ii–iii.2004) (1ɗ EFN), 3 adults obtained (vi–vii.2004) from larva idem collected 12.v.04 (1ɗ, 2Ψ EFN). SPAIN: Sierra de Gredos: Piedralves, Hi. (18.v.1934, coll. C. Koch 2Ψ, PCCC, MSNM); Vigo: (Before 1927, 1Ψ coll. G. C. Champion, BMNH); Malaga: Rd. Ronda – Marbella, 6–20 km SE Ronda, beating dead cut branches of Pinus sp. (18.v.67, coll. M. E. Baechus & B. Levey, 1Ψ, BMNH).
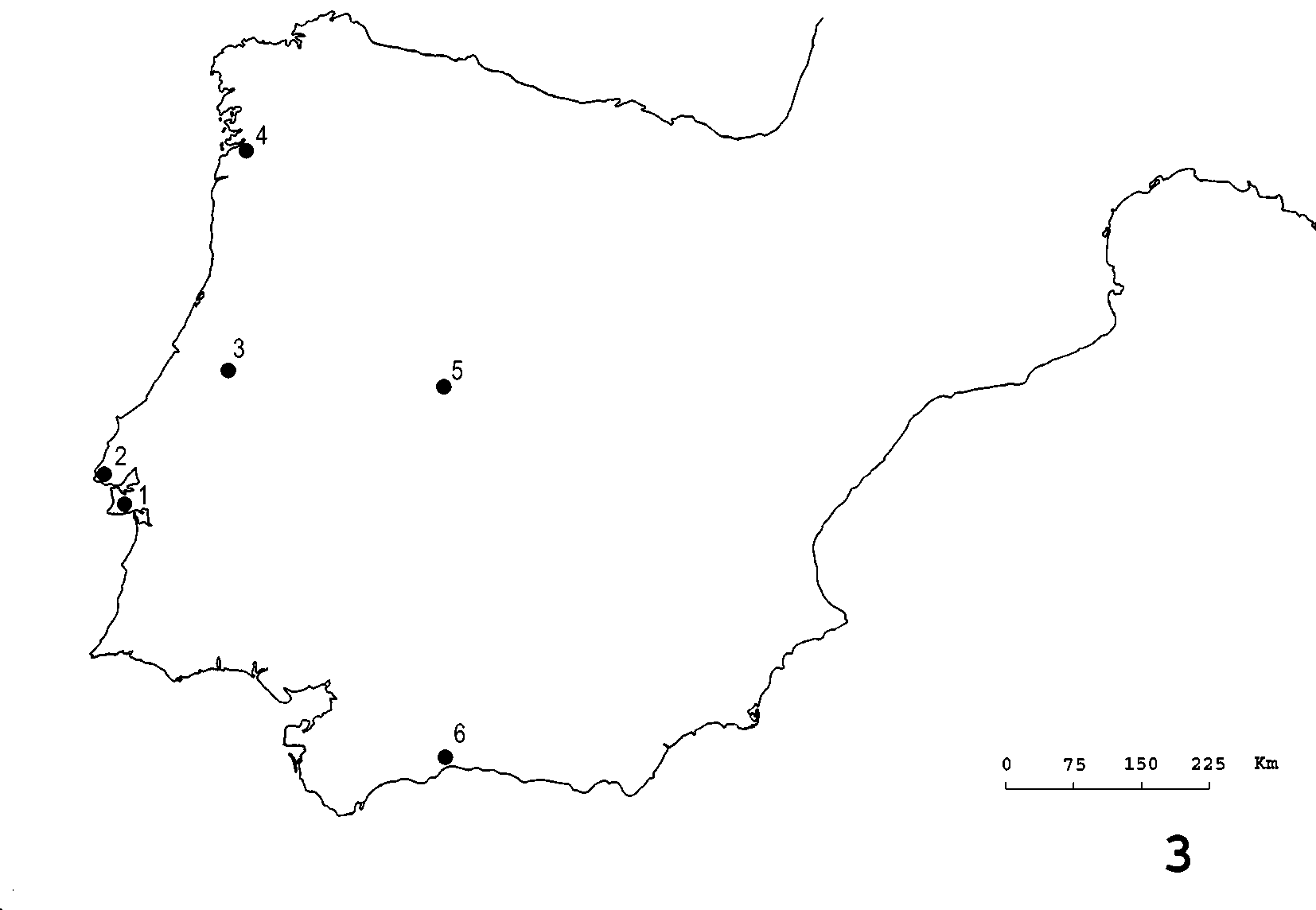
Distribution. Iberorhyzobius rondensis is only known from the Iberian Peninsula, and is widely distributed from the southeast coast of Andalusia up to the northwestern region of Galicia and Portugal ( Fig. 3 View FIGURE 3 ). In Portugal, the species was found at an altitude range of 25 m – 600 m.
Differential diagnosis. Iberorhyzobius can be easily differentiated from both Rhyzobius and Coccidula spp. by several features: absence of prosternal carinae, sparse and fine pubescence, body dimensions and body shape elongated/oval ( Figs. 1–2 View FIGURE 1 View FIGURE 2 ).
Specimens of Coccidula possess a pair of prosternal carinae joined apically (absent in Iberorhyzobius , Fig. 2 View FIGURE 2 ) and body sides almost parallel ( Iberorhyzobius has an oval shape). Additionally, Coccidula exhibits eyes only moderately coarsely faceted ( Iberorhyzobius differs by the eyes strongly coarsely faceted).
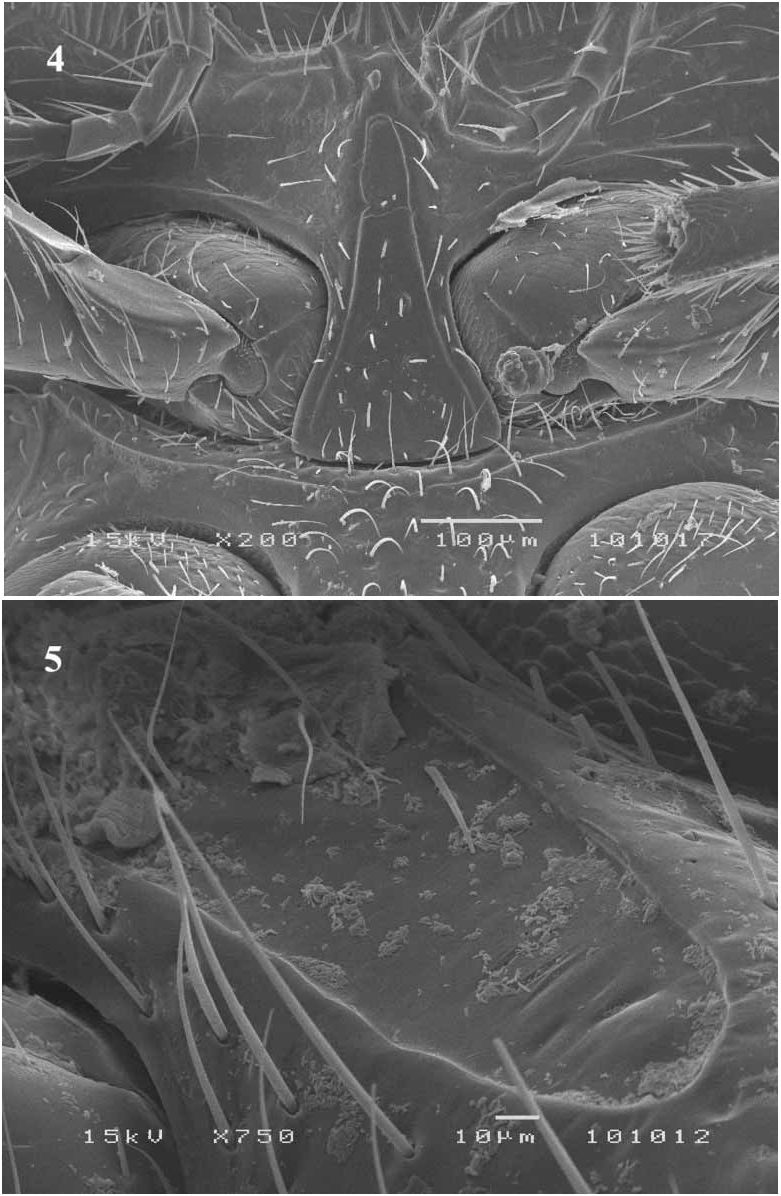
Rhyzobius differs from Iberorhyzobius by the presence of a pair of prosternal carinae ( Figs. 4–5 View FIGURES 4 – 5 ).
Both Rhyzobius and Coccidula are distinguished from Iberorhyzobius by the dorsal dense pubescence. Rhyzobius has ventral and dorsal surfaces of the body covered by yellowish semierect pubescence always distinct (e.g. Gordon, 1994). When observed under the binocular microscope the pubescence of Iberorhyzobius is fine and sparse. Observation by SEM revealed two distinct kinds of pubescence ( Fig. 2 View FIGURE 2 ): i) thinner and abundant and ii) thicker and scattered. The structure of genitalia of I. rondensis easily differentiates it from the other Coccidulini species ( Fig. 6 View FIGURE 6 ) as follows: median lobe elongated, asymmetrical, slightly longer than paramere; sipho elongated and with apex simple; siphonal capsule with outer arm absent. Female genitalia with genital plates very elongated, infundibulum and receptaculum seminis as in others Coccidulini .
Description of 4th instar larva. Body elongate oval, colour brown. Body length 3.6 mm, body width 1.2 mm, head width 0.75 mm, length of front tibiotarsus 0.25 mm, length of middle tibiotarsus 0.30 mm. Head distinctly sclerotized, subquadrate; epicranial stem absent; epicranial arms curved laterally from vertex to three fourths of the head then directed laterally for the last fourth; antennae threesegmented. Prothorax subtrapezoidal with lateral margin crenulate and with a lateral series of chalazae. Meso and metathorax with series of lateral and dorsolateral chalazae. Abdomen 9segmented with series of lateral and dorsolateral strumi covered in each margin by 5–6 chalazae.
Natural history. Larvae were collected on belt traps installed on tree trunks to study the phenology of the maritime pine bast scale, in central and southern Portugal, and were observed to feed on the eggs of this scale insect ( Fig. 5 View FIGURES 4 – 5 ). Larvae showed a kairomonal response to the sex pheromone of the prey, M. feytaudi ( Branco et al. 2006) . Adults are also attracted to the prey sex pheromone, but they display a weaker response. Observations taken from 2000 to 2006, using both tree belt traps and tree arenas, set up with rubber dispensers loaded with the sex pheromone of the prey, showed that larvae were only active from February to MayJune. Adults were observed only in MayJune. These data suggest a univoltine life cycle.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |