
Incadelphys antiquus Marshall & Muizon, 1988
|
publication ID |
https://doi.org/ 10.5252/geodiversitas2022v44a22 |
|
publication LSID |
urn:lsid:zoobank.org:pub:B8864E92-2EC0-4E0A-97F1-BFAE94DC24D8 |
|
DOI |
https://doi.org/10.5281/zenodo.6802477 |
|
persistent identifier |
https://treatment.plazi.org/id/03D2AB06-FFCE-5129-F8EC-2395DEE7E716 |
|
treatment provided by |
Felipe |
|
scientific name |
Incadelphys antiquus Marshall & Muizon, 1988 |
| status |
|
Incadelphys antiquus Marshall & Muizon, 1988
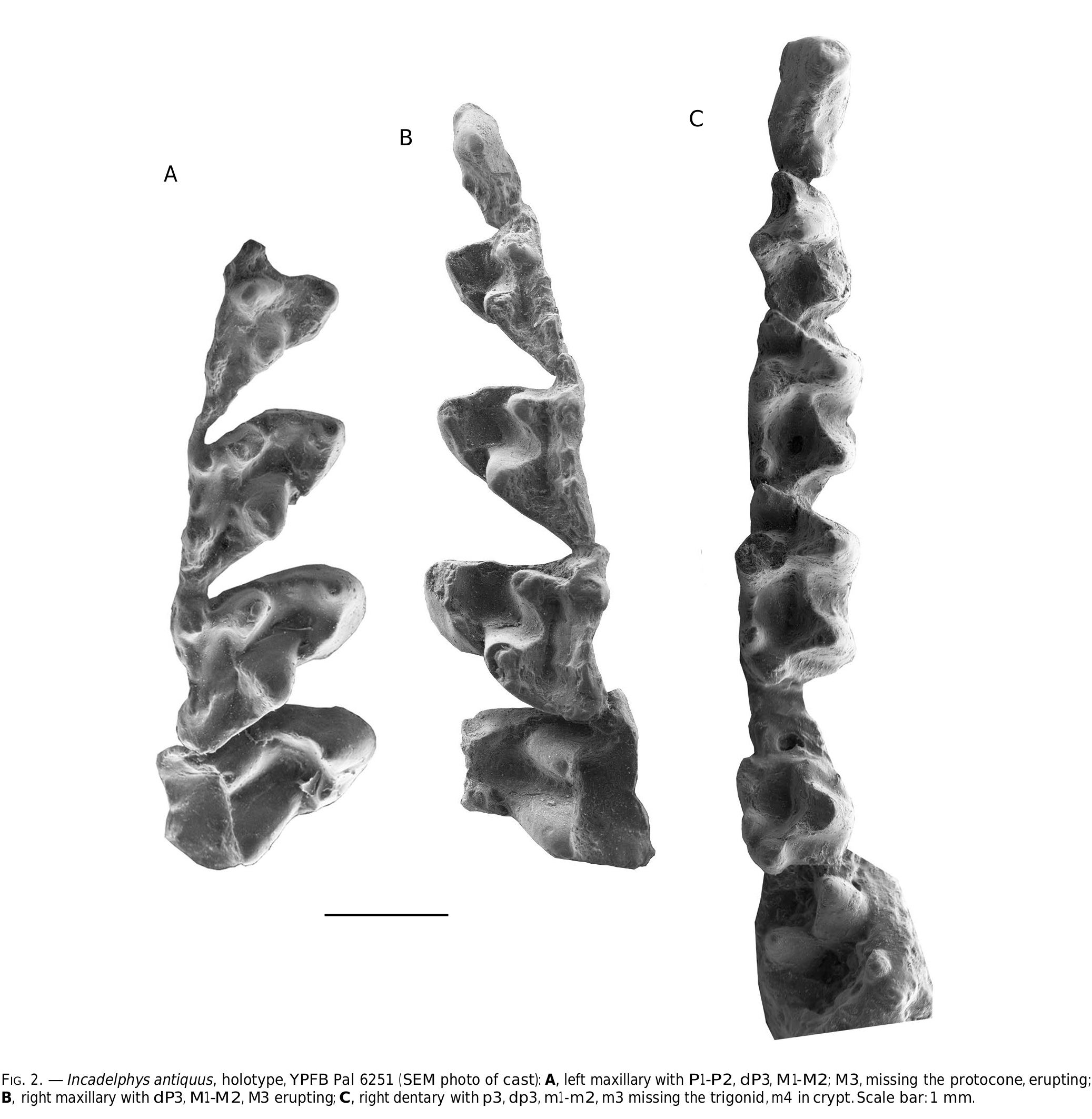
HOLOTYPE. — YPFB Pal 6151 , partial upper and lower jaws of the same juvenile specimen including the left maxilla with base of P1, P2, dP3, unerupted P3, M1-M3 (M3 erupting and missing tip of protocone); the right maxilla with dP3 unerupted P3, M1-M3 (M3 erupting); the right mandibular ramus with p1, p2, dp3, unerupted p3; m1-2, talonid of m3, unerupted m4; the left mandibular ramus with talonid of dp3, m1-2, m3 broken, m4 unerupted ( Fig. 2 View FIG )
EMENDED DIAGNOSIS. — Dental formula I5/i4, C/c, P3/p3; M4/ m4; skull slightly smaller than Pucadelphys but distinctly larger than Szalinia ; approaching the size of the extant didelphid Thylamys , proportions of the rostrum approaching those of Pucadelphys with the apex more slender; palatal vacuities absent.
Incadelphys antiquus differs from Pucadelphyidae in the following features: occurrence of a distinct lacrimal-nasal contact; extremely narrow and blade-like upper premolars, with weaker labial and lingual posterocingula (in pucadelphyids upper premolars are wider with thick labial and lingual posterocingula); upper molar smaller and more gracile; M1 strongly asymmetrical; distolabial angle of M1 conspicuously extended distolingually with angle between labial edge of tooth and postmetacrista varying from 34° to 37° (in pucadelphyids the angle vary from 49° to 62°; mean = 51.4°); protocone mesiodistally shorter; mesiodistal constriction (i.e. shortening) at lingual base of para-metacone more pronounced, especially on M3, (in pucadelphyids constriction is weak to absent); posterolingual inflation of protocone absent or faint (in pucadelphyids posterolingual inflation present); centrocrista weakly V-shaped (in pucadelphyids V-shaped centrocrista is generally conspicuous); anterior stylar shelf on M1 narrower; stylar cusp C absent on M3, absent or small on M1-M2 (in pucadelphyids stylar cusp C is generally present in M1- M3); when present, stylar cusp C much smaller than B and D (in pucadelphyids stylar cusp C is generally subequal to slightly smaller than D); distolabial angle of M1 extending distolabially to a greater extent, with angle between postmetacrista and lingual edge of the tooth smaller; ectoflexus totally absent on M1 (present in pucadelphyids) and shallower on M2-M3 (deep in pucadelphyids); ventral edge of dentary less convex; coronoid process distinctly narrower at apex and not recurved posteriorly (in other words posterior edge of process straight) (in pucadelphyids apex of coronoid process is strongly recurved posteriorly); retromolar space longer (almost as long as m4) (in pucadelphyids it is generally as long as or shorter than talonid of m4); lower molars proportionally narrower; entoconid proportionally larger, slightly higher as compared to hypoconid (in pucadelphyids the entoconid is approximatively as high as the hypoconid).
Incadelphys antiquus differs from Aenigmadelphys archeri in that the paracone is slightly smaller in height and volume than the metacone (reversed in A. archeri ); paraconule subequal to metaconule (in A.archeri paraconule is larger than metaconule); ectoflexus absent on M1 (present and shallow in A. archeri ); anterior stylar shelf narrower than posterior (in A. archeri the anterior stylar shelf is wider than the posterior on the holotype only [an M3], but it is narrower on OMNH 23460, [an M3 lacking the protocone], OMNH 20120 [an M2], and OMNH 22898 [an M1]); stylar cusp B and D subequal in size (in A. archeri stylar cusp B is consistently larger than D); stylar cusp C absent on M3 (in A. archeri it is small but distinct); trigonid lower as compared to talonid; paraconid distinctly smaller than metaconid (in A. archeri paraconid is subequal in size to metaconid or slightly smaller).
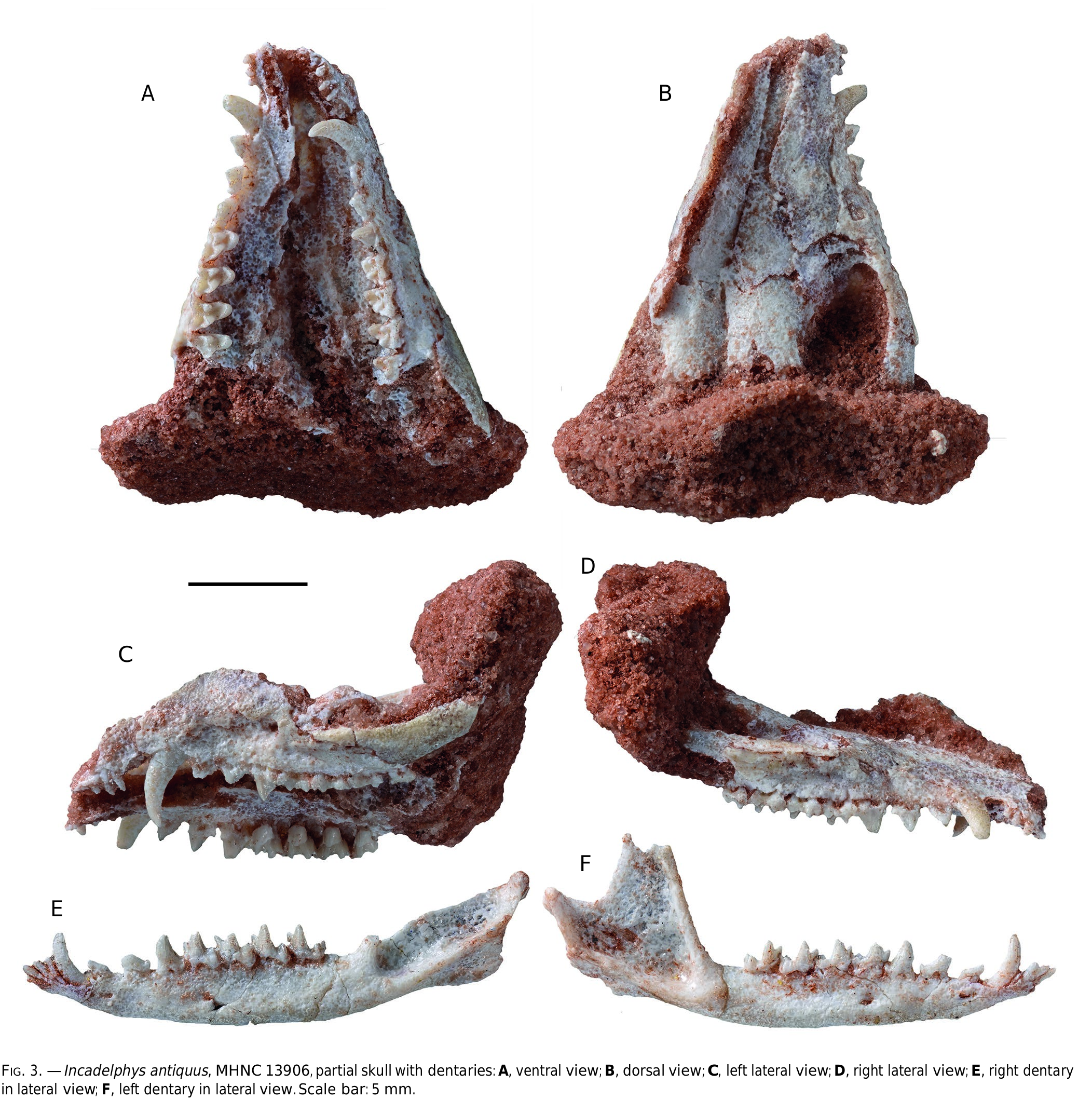
HYPODIGM. — The holotype; MHNC 13906 View Materials , anterior half of a skull including both premaxillae, maxillae, nasals, lacrimals, anterior part of the frontals, anterior part of the jugals, right I3-M4, left I1-M4 (crown of I1-2 broken at base; labial edge of stylar shelf of M3-4 missing); left dentary (missing coronoid process) with i1-m4; right dentary with i2-m4 (labial edge of protoconid of m1- m3 abraded, associated to the skull, the specimen also includes seven caudal vertebrae, three metacarpals and three metatarsals ; MHNC 13947 View Materials , a partial maxilla with M1-M3 ; MHNC 8270 View Materials , a left mandible, with c-m3 and alveoli of m4 (on molars lingual edge of para- and metaconid is scratched) , MHNC 13933 View Materials , a left M2 , MHNC 13935 View Materials , a left M2.
GEOLOGICAL SETTING AND AGE. — All the specimens of Incadelphys antiquus are from beds of the Santa Lucía Formation at Tiupampa and have been discovered in the locality called “the Quarry” by Gayet et al. (1992) and Marshall & Muizon (1995). As discussed by Gelfo et al. (2009), Muizon et al. (2015, 2018), Muizon & Ladevèze (2020), the age of the Tiupampa beds is regarded as early Danian in age (c. 65 Ma) contra Zimicz et al. (2020).
COMPARATIVE DESCRIPTION
The new specimen of Incadelphys antiquus described here is an anterior part of a skull including the upper tooth rows and the palate, and both dentaries ( Fig. 3 View FIG ). The specimen has suffered some distortion and the left maxilla has been displaced dorsally. In spite of this distortion, it seems that the width of the rostrum has not been strongly affected. The specimen includes the premaxillae, the maxillae, the lacrimals, the nasals, the anterior part of the frontals and jugals. The dentaries are almost complete with all their teeth. However, during collection of the specimen, the lateral side of the left tooth rows have been damaged; on upper teeth, the posterolabial angle of M3 and the labial edge of the stylar shelf of M4 have been destroyed. On the lower teeth, the labial edge of the protoconid and hypoconid of m1-m3 have been scratched during the collection of the specimen.
Dentition
The dental formula is the plesiomorphic pattern for metatherians: I5/i4, C/c, P3/p3, M4/m4 (Kielan-Jaworowska et al. 2004). The upper dentition will be described first, followed by the lower dentition.
Upper dentition
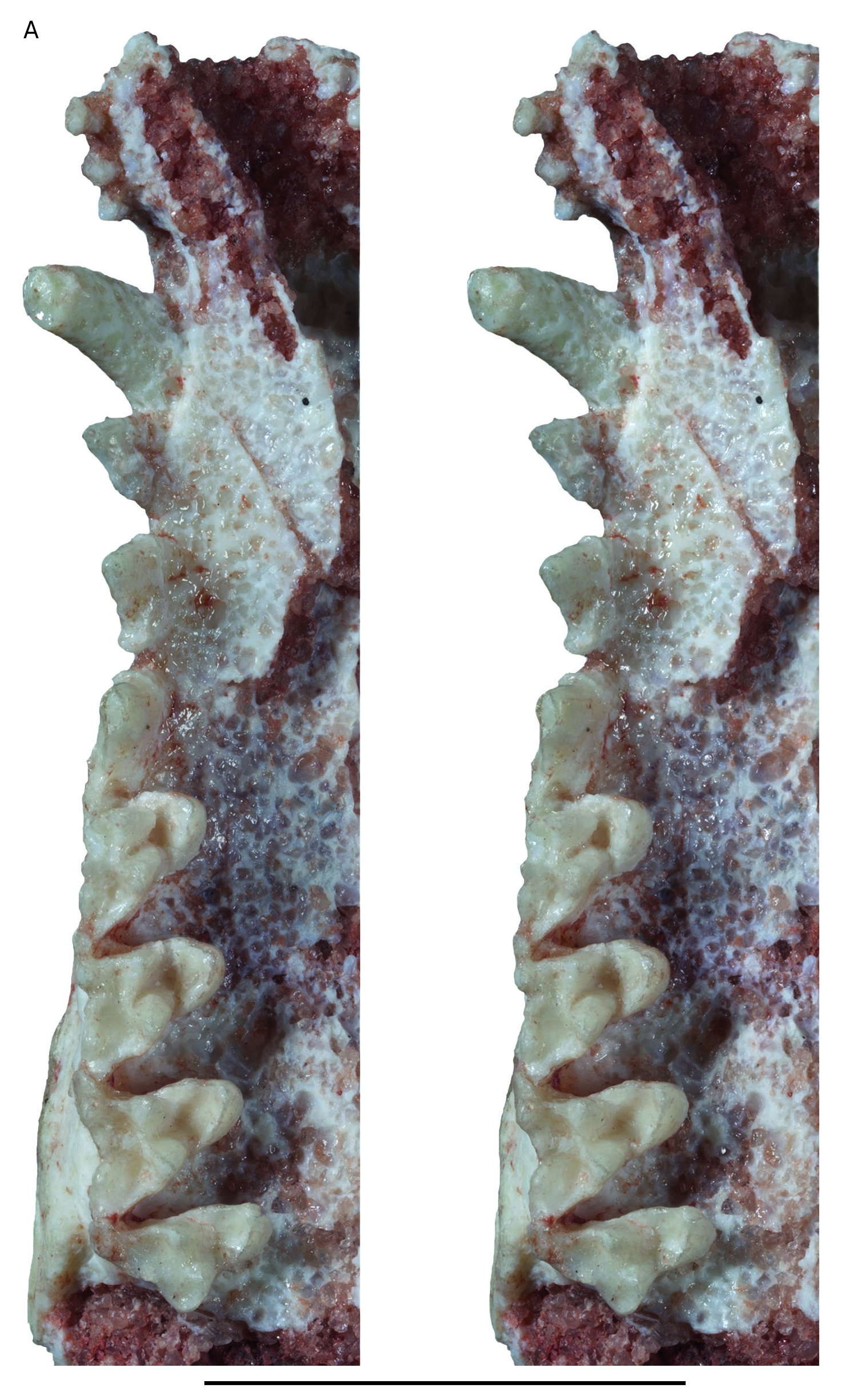
Upper incisors ( Figs 4 View FIG ; 5 View FIG ). the crowns of I1 and I2 are missing and the roots of these teeth are preserved only on the left premaxilla. I1 has a slightly smaller diameter than I2. This condition contrasts with that of extant didelphids and Andinodelphys , in which I1 is distinctly larger than I2. In contrast, it resembles the condition observed in the only Pucadelphys specimens that preserve the I1s (YPFB Pal 6105, the holotype, and MHNC 8378, referred to females by Ladevèze et al. 2011). A small diastema is present between I1 and I2. The diastema is approximately as long as the diameter of I1. A similar condition is present in some specimens of Pucadelphys (MHNC 8378) and in didelphids but is absent in Andinodelphys . The crowns of the other incisors are preserved. They are subequal in size to I2, with I5 being slightly smaller. Their crown is peg-like. It is slightly compressed labiolingually on I3 and I4 and roughly conical on I5.
Upper Canine ( Figs 4 View FIG ; 5 View FIG ). The upper canine is large and sharp and is approximately three times as high as the P3. It is larger than in Marmosa , in which it is twice as high as P3, and Thylamys , in which it is less than twice the high of P3. It is similar in height to the canines of Didelphis and Caluromys , in which they are approximately three times (or more) as high as the P3. Among the Tiupampa metatherians, the canine of Incadelphys is similar in relative height to those of Andinodelphys and specimens interpreted by Ladevèze et al. (2011) as males of Pucadelphys (e.g., MHNC 8266, 8377, 8382). In contrast, it is clearly higher than the canines of the specimens referred to females of Pucadelphys by these authors (e.g., MHNC 8376, 8378), in which the canine height is less than twice that of P3. As in all didelphids and pucadelphyids, the upper canines of Incadelphys are transversely compressed being approximately twice as long as wide. As in didelphids and pucadelphyids, they are strongly curved posteriorly. The upper canine of Incadelphys is slightly procumbent as indicated by the position of the apex of the tooth, which is ventral to the anterior edge of the crown base.
Upper premolars ( Figs 4 View FIG ; 5 View FIG ). The three premolars are doublerooted and distinctly increase in size from P1 to P3. As observed on the right side of the skull, the increase in size is progressive. In other words, the increase in size between P1 and P2 is similar to that between P2 and P3. This condition differs from that observed in Andinodelphys , in which a great increase in size is observed between P1 and P2 and a smaller increase exists between P2 and P3. A small diastema is present between P1 and P2, and a smaller one between P2 and P3. P1 and P2 are extremely narrow transversely and blade-like, whereas P3 is slightly wider ( Table 1 View TABLE ). The blade-like morphology of the P1-P2 is not observed in any of the other Tiupampa metatherians. As shown on Table 2 View TABLE the ratio W/L for the P2 of Incadelphys is 0.31 (three measurements), 0.45 for Andinodelphys (seven measurements), and for Pucadelphys 0.56 (ten measurements). Therefore, the relative width (as compared to length) of the P2 of Incadelphys is 31% smaller than in Andinodelphys and 45% smaller than in Pucadelphys . Given these results, the blade-like morphology of the anterior upper premolars of Incadelphys is regarded as a significant characteristic of the genus. P1 is triangular in lateral view. It is slightly asymmetrical, with its apex located below the anterior root. It is less asymmetrical than in Andinodelphys , in which the apex of P1 is ventral to the posterior edge of the posterior root. The P1 of Incadelphys has no posterior accessory cusp, nor cingulum. Its anterior root contacts the posterior edge of the canine, but it is not closely appressed against it, thus differing from the condition of Andinodelphys . P2 is less asymmetrical than P1 and its apex is ventral to the inter-alveolar septum. Similarly to P1, it is as long as high. Its anterior and posterior edges are straight. The tooth bears a small cingulum at the anterior edge of the crown, which forms a hint of an anterior basal cuspule. The cingulum extends on the lingual aspect of the crown, below the anterior root. Posteriorly a conspicuous basal cusp is present. P3 is more robust and more inflated than P2. Its anterior edge is slightly convex and its posterior edge slightly concave. The apex of the crown is located below the anterior edge of the posterior root. The anterior border of the crown is rounded, whereas the posterior edge becomes thinner, sharp, and crest-like. The lateral edges of the crown, in this region, are distinctly concave. The anterior cingulum is strong (as compared to that of P2). It extends on the mesiolabial and mesiolingual angles of the crown but remains at the level of the anterior root. Posteriorly, the basal accessory cusp is well developed. It imbricates in the mesiolingual angle of M1 with the stylar cusp A. Labial and lingual to the basal cusp are small cingular shelves.
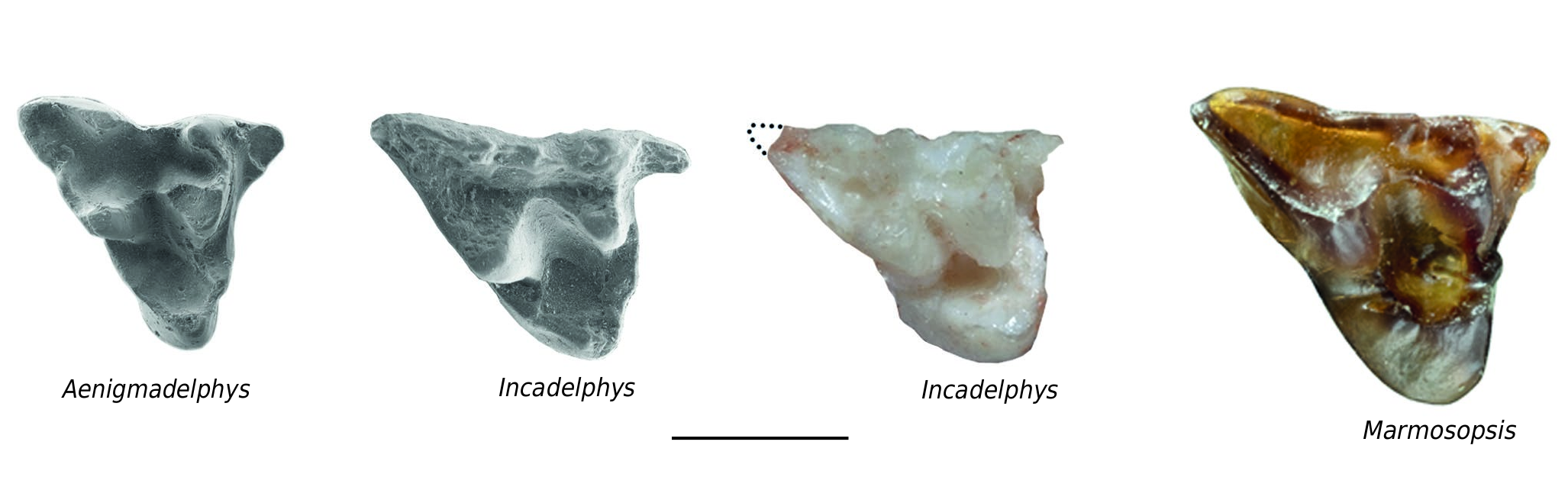
Upper molars ( Figs 4 View FIG ; 5 View FIG ). The description of the upper molars will consider the five available specimens: the holotype YPFB Pal 6251, which includes both maxillae and mandibles of a subadult individual (i.e. upper M4 are missing and m4 are unerupted); MHNC 13906 (which preserves complete maxillae and mandibles and 13931 a partial maxilla with M1-3). M1 is relatively different from the other molars and will be treated first. M2-M3, which have a more typical morphology as compared to most of the other Tiupampan metatherians will be described jointly. M4, which strongly differs from the preceding molar, as in most metatherians, will be studied next. M1 is a distinctive tooth of Incadelphys in the distolabial extension of its metastylar angle. As a consequence, the labial edge of M1 is much longer than on the other molars ( Table 1 View TABLE ). The anterior edge is shorter than on the posterior molars, and the tooth is longer than wide (1.56 mm vs 1.54mm; mean of five measurements, seeTable 1), a condition that contrasts with that of Andinodelphys , Pucadelphys , and Mizquedelphys . A condition of the M1 similar to that of Incadelphys is observed in the Campanian genus Aenigmadelphys of Utah. Table 3 View TABLE compares the angle between the line joining stylar cusps D and E and the postmetacrista. Because the postmetacrista is often curved, and because the posterolabial part of the crista is relatively straight, the latter has been used for the measurement. As observed inTable 2, the mean of the angle obtained for pucadelphyids (53.2°) is approximately 48% and 32% greater than the angle of Incadelphys and Aenigmadelphys respectively. Because of this difficulty of measurement, the value of the angle obtained is certainly somewhat imprecise. However, because of the great difference observed, this result is probably significant. As a consequence of the posterolabial extension of the M1, its mesial edge is distinctly shorter than its labial edge. A similar condition is also observed in Aenigmadelphys . Marmosopsis from the early Eocene of Itaboraí, clearly ranges close to Incadelphys , whereas Monodelphis and Thylamys are closer to pucadelphyids than to Incadelphys and Aenigmadelphys .
The protocone of M1 is relatively massive, being mesiodistally longer than wide on the three specimens. It is roughly symmetrical mesiodistally and does not present the distolingual inflation observed in didelphids and pucadelphyids. The mesial and distal bases of the protocone are smooth and bear no cingulum. Para- and metaconules are well-developed (almost as large as stylar cusps B and D). From the paraconule, a conspicuous paracingulum (i.e. labial extension of the preparaconular crista) extends up to stylar cusp A. Distally, the metaconule abuts the distal base of metacone but no metacingulum (i.e. labial extension of the postmetaconular crista) is present. The paracone is slightly smaller in height and volume than the metacone. Both cusps are widely separated at base, as in didelphids and pucadelphyids. A deep trigon basin is bordered by the distolingual aspect of the paracone, the mesiolingual aspect of the metacone and the labial aspect of the protocone. The lingual aspect of the para- and metacone is strongly convex, whereas their labial aspect is flat to slightly concave. As a consequence, the cusps are triangular in section and the pre- and post- paracristae and pre- and post-metacristae are shifted labially, forming the lingual wall of the stylar shelf. The junction of the postparacrista and premetacrista (i.e. the median point of the centrocrista) is displaced labially, and the centrocrista is slightly V-shaped in occlusal view. This condition, which is present in Aenigmadelphys , didelphids, and pucadelphyids, differs from that observed, for instance, in Kokopellia , alphadontids, peradectids, and sparassodonts, in which the labial and lingual aspects of the para- and metacone are markedly convex (generally slightly less convex labially than lingually), the pre- and post- paracristae and pre-and post-metacristae are in a median position relative to the para- and metacone and therefore the centrocrista is straight. The stylar shelf of Incadelphys is narrow, being almost absent anteriorly since the stylar cusp B is almost connate to the paracone to which it is connected by a very short preparacrista. The stylar cusp A is smaller than B but conspicuous. It surrounds posterolabially and imbricates with the posterobasal cusp of P3. Stylar cusp B and D are large and subequal in size, but variation exists. In MHNC 13906, StB is slightly smaller than StD, whereas in MHNC 13931 StB is slightly larger than StD; these cusps are subequal in the holotype. Stylar cusp B is conical, whereas stylar cusp D is transversely compressed. Between StB and StD, a small stylar cusp C is present in the holotype and MHNC 13906. Stylar cusp C is lacking in MHNC 13931. Stylar cusp E is indistinguishable. The labial edge of M1 is straight and features no ectoflexus.
M2 and M3 are conspicuously wider than long. They differ from M 1 in the protocone, which is approximately as long as wide on M2 and clearly wider than long on M3. The stylar shelf is wider than on M1, especially its anterior part. As a consequence, the stylar cusp B is well separated from the paracone, and the preparacrista is as long as the postparacrista on M2 and slightly longer on M3. The preparacrista contacts stylar cusp B on its anterior edge, almost between cusp B and A on MHNC 13906. Stylar cusp C is small to absent and stylar cusp D is well developed but smaller than B. On the labial edge of M2 is a small ectoflexus (rather a notch) between stylar cusps B and C. On M3, the ectoflexus is deeper and located between stylar cusps C and D. From M1 to M3, at the distolabial corner of the tooth, the angle between the labial edge and the postmetacrista increases and, as a consequence, the postmetacrista is more transverse posteriorly. M2-M3 of Incadelphys strongly resemble those of Aenigmadelphys , from which they differ however in the latter being transversely wider and mesiodistally shorter (see comparison below p. 000). Furthermore, the M2-M3 of Aenigmadelphys have a slightly wider stylar shelf.
On M4, the protocone is shorter mesiodistally than on the anterior molars and the metacone is greatly reduced compared to that of the anterior molars, being significantly smaller than the paracone. The anterior stylar shelf is wider and the preparacrista is longer than on M2-M3. The posterior stylar shelf is very narrow but still present labial to the metacone. The M4 of Incadelphys differs from that of Aenigmadelphys in the latter being mesiodistally shorter, transversely wider with a longer preparacrista, and in the posterior stylar shelf being virtually absent.
Lower dentition
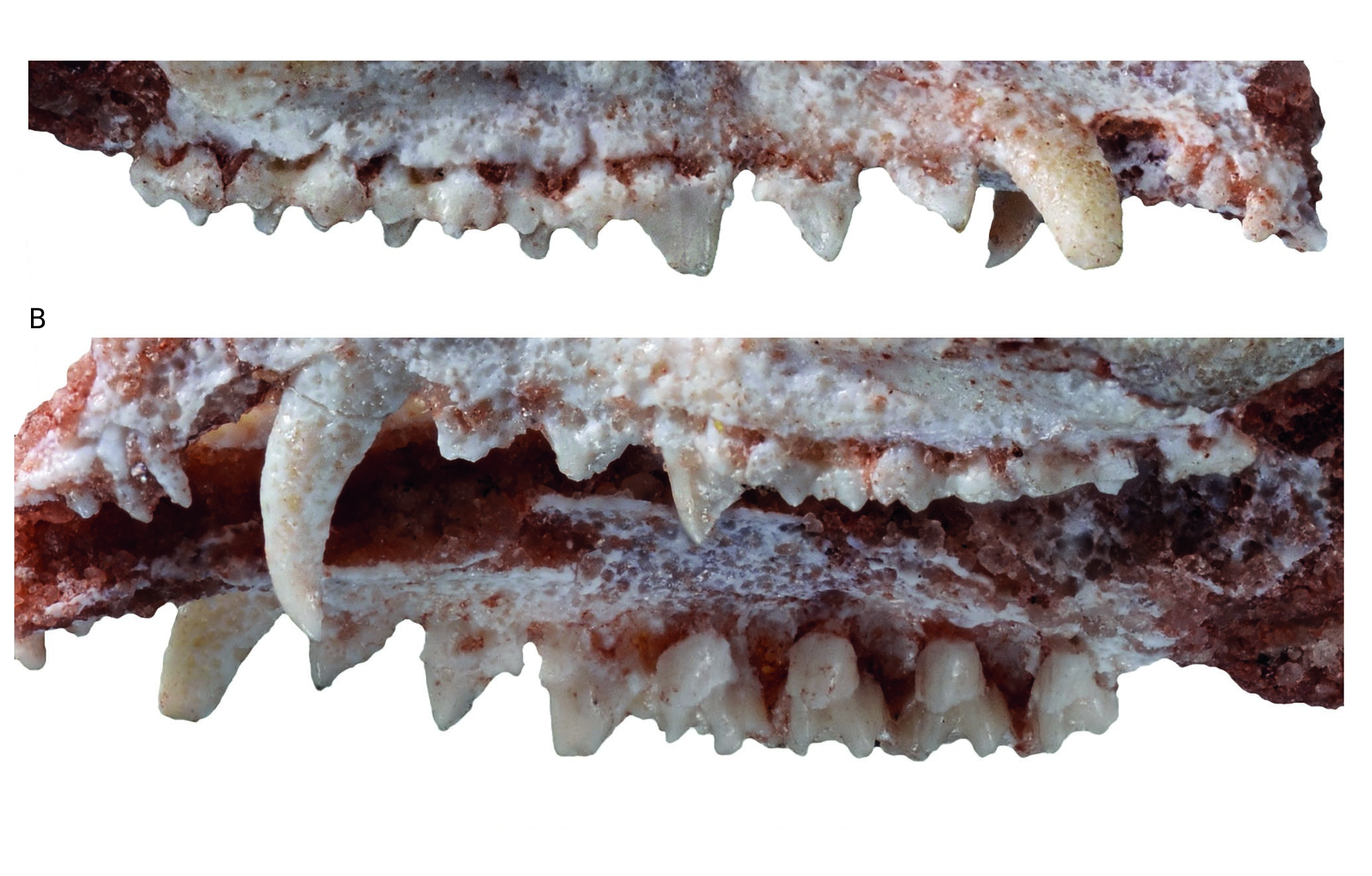
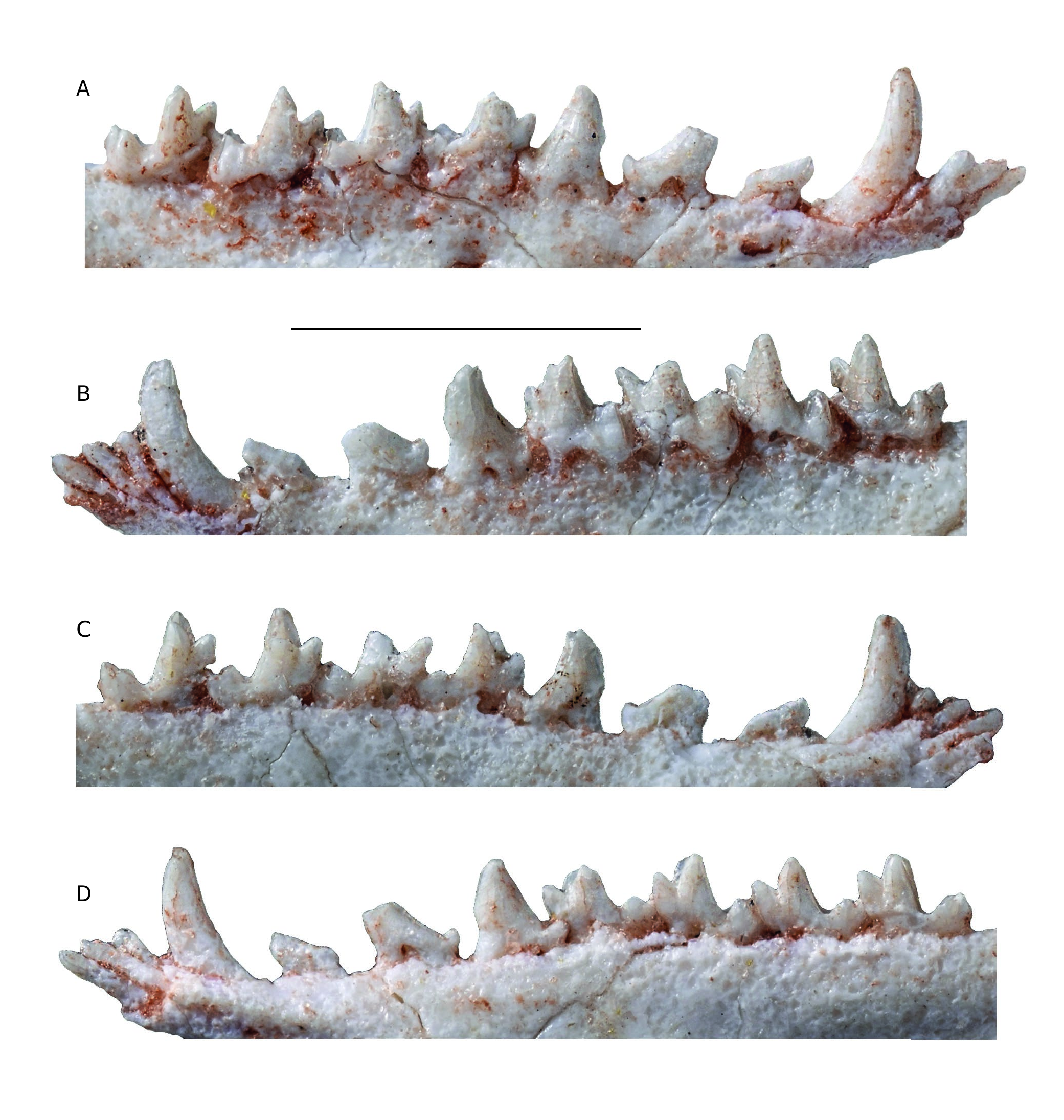
Lower incisors ( Figs 6 View FIG ; 7 View FIG ). The four incisors are preserved on the left dentary, but i1 is missing part of its crown. In the description below, we follow the interpretation of Hershkovitz (1982, 1995), that the four lower incisors of metatherians are serially homologous to i2-i5. On the right dentary i2-i4 are preserved but i5 is missing its crown. In size, the crown of i2 is smaller than that of i3 but larger than i4; i5 is the smallest of the four incisors. The crowns are peg-like but slightly spatulate, being wider than high. The apex of the crown of i2 is roughly semicircular in labial or lingual view; that of i3 is lanceolate with a carina on its lingual aspect; that of i4 is similar to i1 but smaller and that of i5 is also somewhat lanceolate. The i3 is distinctly staggered as indicated by the buttress visible on the anterolabial edge of the dentary just ventral to the labial aspect of the tooth and by the posterior shift of its root observable lingually on MHNC 13906.
Lower canine ( Figs 6 View FIG ; 7 View FIG ). The lower canine is a large tooth, although consistently smaller than the upper canine. It is pointed at its apex and curved (from the alveolar border) dorsally but not posteriorly. In other words, the apex of the tooth does not overhang the distal edge of the crown, which is procumbent ( Fig. 7 View FIG ). The canine is slightly recurved lingually, this feature being more pronounced on the right dentary. Its anterior edge is closely appressed against the posterior edge of i5.
Lower premolars ( Figs 6 View FIG ; 7 View FIG ). The lower premolars are well preserved on MHNC 13906. From p1 to p3 the lower premolars of Incadelphys conspicuously shift from a mesially tilted to an upright position. The p1 is very small and procumbent, being strongly asymmetrical in lateral view. Its apex is strongly shifted mesially and overhangs the anterior edge of the anterior root of the tooth. As a consequence, the posterior edge of the tooth is very long and strongly oblique. It is straight and bears no cuspule at its distoventral end. The anterior edge of the crown is markedly convex and extends mesially over the posterior edge of the canine. This highly asymmetrical morphology of the p1 of Incadelphys resembles that observed in Szalinia , in Andinodelphys , in most specimens of Pucadelphys , and in Marmosposis. It is also present in Monodelphis and Marmosa . A small (approximately half the length of p1) diastema separates p1 and p2; a diastema half as long separates p2 from p3. The p2 is larger than p1 being intermediate in size between the latter and p3. It is also strongly shifted anteriorly but less procumbent than p1. Because its apex is missing on both sides, its position relative to the root cannot be evaluated precisely. It is clear, however, that is was overhanging the anterior root of the tooth. The anterior edge is strongly convex and projects anterior to the anterior edge of the anterior root. The posterior edge is straight and bears a sharp and pointed accessory cusp at its distoventral end. The p3 is robust and roughly as high as long. It is subvertical, not procumbent, and its apex overhangs the middle region of the anterior root. The anterior edge of the tooth is subvertical but still convex anteriorly and projects above the anterior edge of the anterior root. The posterior edge of p3 is longer than the anterior but relatively less, as compared to anterior premolars. It is slightly concave dorsally and bears a large accessory cusp at its base. None of the premolars bears any kind of cingulum.
Lower molars ( Figs 6 View FIG ; 7 View FIG ). The lower molars slightly increase in length distally from m1 to m3. The m4 is slightly shorter than the preceding tooth. The length of the trigonid decreases posteriorly from m1 to m4. On the four molars, the trigonid is more than two times as high as the talonid. The trigonid of m1 is relatively long with a paraconid set mesiolabially (as compared to the other molars) so that the angle between the paracristid and protocristid is more open lingually. As a consequence, the trigonid of m1 is distinctly longer than wide, while it is approximately as wide as long on m3-4 (measure not available on m2). The protoconid is the largest cusp of the trigonid; it is triangular in section, while the metaconid is roughly ovoid and less voluminous. In lingual view, the metaconid is subvertical and slightly decreases in length distally; its posterior edge is sloping on m1 and becomes subvertical on m4. The paraconid is the smallest cusp of the trigonid. In lingual view, it is consistently tilted mesially, a condition that reduces on posterior molars.
Paraconid and metaconid are connate at base, and the vallid between them is higher than half of the metaconid height (measured from the lingual alveolar border). Because of this condition, the trigonid basin is elevated, well-excavated, and well-circumscribed lingually at least on m2-4. On these molars, the deepest point of the basin is at the level of, or lower than, the uppermost point of the paraconid-metaconid vallid. This condition is similar to that observed in Pucadelphys but differs distinctly from that observed in the Tiupampa sparassodonts, Mayulestes and Allqokirus . In these taxa, the paraconid and metaconid are broadly separated and the vallid between them is wider and extends almost as far as the base of the crown lingually. As a consequence, the trigonid basin of these sparassodonts is broadly open lingually and is more a slope on the lingual side of the protoconid than a true depression. The paraconid of Incadelphys is triangular in section and its mesiolingual angle bear a moderately salient paraconid ridge. Labial to this ridge is a shallow hypoconulid notch. The paracristid and protocristid are sharp but bear no carnassial notch. The median point of the cristids (the point of contact of the cusps) is slightly lower on the paracristid than on the protocristid, this condition being more marked on the anterior molars. The paracristid is strongly oblique relative to the axis of the tooth row, whereas the protocristid is slightly oblique on m1-m2 and distinctly transverse on m3-m4. The angle of the paracristid with the axis of the tooth row increases from m1 to m4.
The talonid is distinctly basined. On m1-3, the hypoconid is the largest cusp in height and volume; the entoconid and the hypoconulid are subequal in height and volume. On m4, the hypoconulid is the highest cusp of the talonid and is almost as voluminous as the hypoconid. It is consistently larger than the entoconid. The hypoconid is large and occupies approximately half of the talonid volume. It is conspicuously larger than the other cusps of the talonid. In occusal view, it is triangular to V-shaped in appearance. At the mesial edge of the hypoconid, the cristid obliqua is well-developed and sharp. It extends mesiolingually and connects to the trigonid at the distolingual edge of the protoconid (i.e. slightly labial to the protocristid notch). On the distolingual edge of the hypoconid, a strong posthypocristid connects to the hypoconulid. It is distolabially concave and deeply notched between the two cusps. The posthypocristid notch is located approximately at midline of the talonid. At the distolingual angle of the talonid, the hypoconulid and entoconid are distinctly connate. The hypoconulid is lingual to the midline of the talonid. The entoconid is larger than the hypoconulid on m1-m2, subequal to hypoconulid on m3, and smaller on m4. On its mesial edge, it bears, a marked entocristid, which connects the distal base of the metaconid. Therefore, the talonid is relatively well enclosed lingually. On m4, the hypoconulid is enlarged and elevated above the other talonid cusps. The entoconid is reduced and partially fused at the base of the hypoconulid. A well-developed precingulid is present at the mesial base of the protoconid and extends on the mesiolabial base of the paraconid. A thick postcingulid is present on distal edge of m1-3 as a shelf extending ventrolabially from the apex of the hypoconulid to the distal edge of the hypoconid as far as its labial side. A postcingulid is absent on m4.
Bony skull
General features. Because the skull of Incadelphys antiquus described here is incomplete, little can be described of its general features.However, the presence of diastemata between the upper and lower premolars is an indication of a relatively elongated rostrum, as observed in the male specimens of Pucadelphys ( Ladevèze et al. 2011) ; the rostrum of I. antiquus was probably also approaching the condition observed in the extant species of Thylamys , but it was not as long, relatively, as that of Andinodelphys . Furthermore, in spite of the rostrum distortion and displacement of the maxillae and premaxillae, it is possible to observe that the apex of the snout was slender and narrow, as in Pucadelphys and Thylamys .
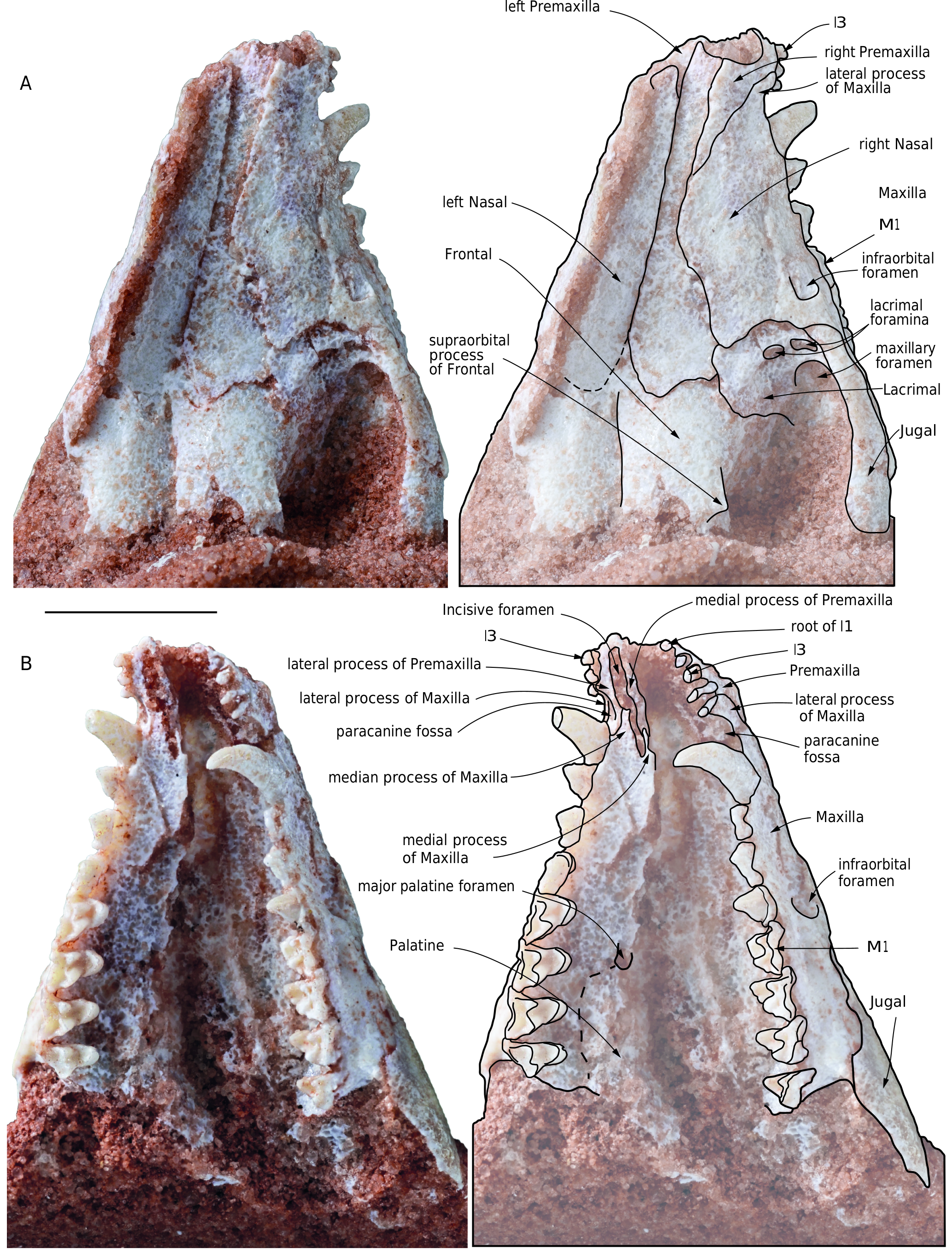
Premaxilla ( Figs 8 View FIG ; 9 View FIG ). In lateral view the premaxilla presents a long and narrow posterodorsal process (the facial process), which is wedged between the nasal and the maxilla. The apex of the process extends posteriorly as far as the posterior third of the parallel-sided anterior portion of the nasal. On the lateral aspect of the rostrum, the premaxilla-maxilla suture is roughly straight. Anteroventrally the suture reaches the alveolar border between I4 and I5. In other words, a small anterolateral process of the maxilla covers the premaxilla in the area of the lateral alveolar border of I5 (see below). The dorsal edge of the premaxilla forms a right angle at the junction of the part that articulates with the nasal and the lateral border of the narial opening. In lateral view, the premaxillae are consistently bent ventrally, to a greater extent than in any other extant didelphids and in pucadelphyids. This is especially clear on the right side of the specimen, which does not show cracks or distortion. On the left side a subvertical crack is observed anterior to the canine but apparently with no (or little) ventral displacement of the premaxilla. A similar condition is present in Allqokirus ( Muizon et al. 2018) , although the taxa do not appear to be closely related. In dorsal view, the anterior edge of the two premaxillae have a strongly convex shape (parabolic).The two premaxillae form approximately the portion of the palate anterior to the anterior alveolar border of the canines, with a small contribution of the anterolateral processes of the maxillae. On the palate, the premaxillae are pierced by the anterior half of the elongated incisive foramina (only observable on the right side). The ventral view of the premaxilla shows two posteriorly oriented branches, which form the lateral and medial edges of the incisive foramina. These branches articulate posteriorly with the maxilla on the palate. The lateral branch of the premaxilla, which borders the incisive foramen laterally, has approximately the same width from I1 to the canine. It is approximately as wide as the incisive foramen. Its posterior extremity is located between the anterolateral and the anteromedian processes of the maxilla (the latter being located between the anteromedial and anterolateral processes), (fig. 8B). The posterolateral branch of the premaxilla bears the medial two thirds of the paracanine fossa for the lower canine, which is located just anterior to the upper canine. This branch is rounded posteriorly and contacts the anteromedial edge of the alveolus of the canine. In the anterior region of the paracanine fossa is the alveolus of the I5, while the fossa extends anteriorly as a narrow sulcus on the medial edge of the alveolar border of I5. In the posterior region of the paracanine fossa, the premaxilla-maxilla suture is V-shaped anteriorly. It enters the palate at the level of the posterolabial alveolar border of I5 (between the apex of the anterolateral process of the maxilla and the premaxilla), runs posteromedially from the lateral aspect of the rostrum, almost reaches the anteromedial edge of the canine alveolus, makes a V-turn, and extends further anteromedially as far as approximately the posterior half of the lateral edge of the incisive foramen. The medial branch of the premaxilla forms the medial edge of the incisive foramen for approximately 80% of its length anteriorly. This medial branch articulates posteriorly with a small anteromedial process of the maxilla and both structures slightly overlap anteroposteriorly. The medial branch is narrower than the lateral one.
The incisive foramen is anteroposteriorly elongated and narrow. It is approximately ten times narrower than long. It extends from a point medial to I3 anteriorly to a point medial to the posterior edge of the alveolus of the canine posteriorly.
Maxilla ( Figs 8 View FIG ; 9 View FIG ). The maxilla forms the lateral wall of the rostrum between the posterodorsal process of the premaxilla and the lacrimal (which represents the facial process). Ventrally, it forms most of the palate between the premaxilla anteriorly and the palatine posteriorly (which represents the palatal process). Anteriorly, the maxilla presents an anterolateral process, which forms the lateral wall of the paracanine fossa between I5 and the canine. Its medial edge is excavated and forms the lateral third part of the fossa. The anterolateral process of the maxilla overlaps the posterolateral edge of the premaxilla to a point labial to I5. Posterodorsally, the facial process of the maxilla forms a broad triangular process, which articulates dorsally with the nasal and ventrally with the lacrimal. It has no suture with the frontal and therefore a distinct lacrimal-nasal suture is present. From the apex of the posterior process, the maxilla-lacrimal suture runs in an anterior direction and turns anteroventrally along the anterior rim of the orbit to contact the jugal at the lower edge of the orbit, just anteroventral to the ventral lacrimal foramen. This triple point, which connects the maxilla, the lacrimal and the jugal, is dorsal to the posterior edge of M1. Posterior to this point, the maxilla has a long suture with the jugal, which extends posteriorly along the ventral edge of the jugal on the lateral face of the skull. At the level of the anterior end of the orbitotemporal fossa, the suture passes on the medial side of the zygomatic arch.
On the lateral aspect of the rostrum is a small anterior foramen of the infraorbital canal. It opens (dorsal to posterior edge of P3 and is at the level of the ventral lacrimal foramen dorsoventrally. Because of the distortion of the specimen it is not possible to evaluate the proportions of this foramen. The infraorbital canal transmits the infraorbital nerve (a branch of the V2) and the infraorbital artery, which innervate and supplies blood to the face respectively.
Anterodorsal to the alveolar border of the canine, the maxilla has a narrow anterior lateral process which forms the lateral edge of the paracanine fossa. The anterior end of the process contact the posterolabial angle of I4 ( Fig. 9 View FIG ).
On the lateral aspect of the rostrum, ventral to the maxillajugal suture and dorsal to the alveolar border of M1-M3, is an elongated fossa which extends from a point dorsal to posterior edge of P3 to anterior edge of M3 ( Fig. 9 View FIG ). A similar fossa has been observed in Allqokirus australis ( Muizon et al. 2018) and Andinodelphys cochabambensis ( Muizon & Ladevèze 2020) and has been interpreted as the origin of the levator labii superioris muscle. Such a fossa is also present in Mayulestes and Pucadelphys . In extant didelphids, the fossa for the zygomaticus and levator labii superioris is generally deep and well developed on the anterolateral region of the jugal but does not excavate the maxilla anterior to the jugal-maxilla suture, although the muscles also originate in part on the maxilla ( Turnbull 1970). Because the origin of the levator labii superioris in Didelphis is ventral to that of the zygomaticus, it is likely that the fossa observed in Incadelphys and the other Tiupampa metatherians corresponds to the origin of the levator labii superioris, while the zygomaticus probably originated on the jugal as in Didelphis , although no fossa for this muscle is observed on the jugal of Incadelphys .
Ventrally, the maxilla forms most of the palate from the incisive foramen anteriorly to the maxilla-palatine suture posteriorly. Although in MHNC 13906 the palatal processes of the maxillae are badly crushed, some observations can be made. Anteriorly on the right maxilla the posterior end of the incisive foramen is preserved. In this region, the maxilla has a thin medial process which borders the incisive foramen posteromedially on its posterior fifth and joins the medial process of the premaxilla anteriorly. The posterolateral border of the incisive foramen is formed by the median process of the maxilla, which is much thicker than the medial process and borders the posterior third of the lateral edge of the incisive foramen. The posterior extension of the incisive foramen up to the posterior edge of the canine as observed in MHNC 13906 is absent in the other Tiupampa metatherians (e.g., Andinodelphys , Pucadelphys , Mayulestes ) and in the extent didelphids.
The left palatal process is less damaged than the right one. No palatal vacuity is observed but, at the level of the posterior edge of M2, a large major palatine foramen is present. The anteroposterior position of the foramen is similar to that observed in Andinodelphys and Pucadelphys . The major palatine foramen transmits the major palatine artery and nerve to the ventral region of the secondary palate. On the posterolateral region of the palate, the maxilla meets the minor palatine foramen.
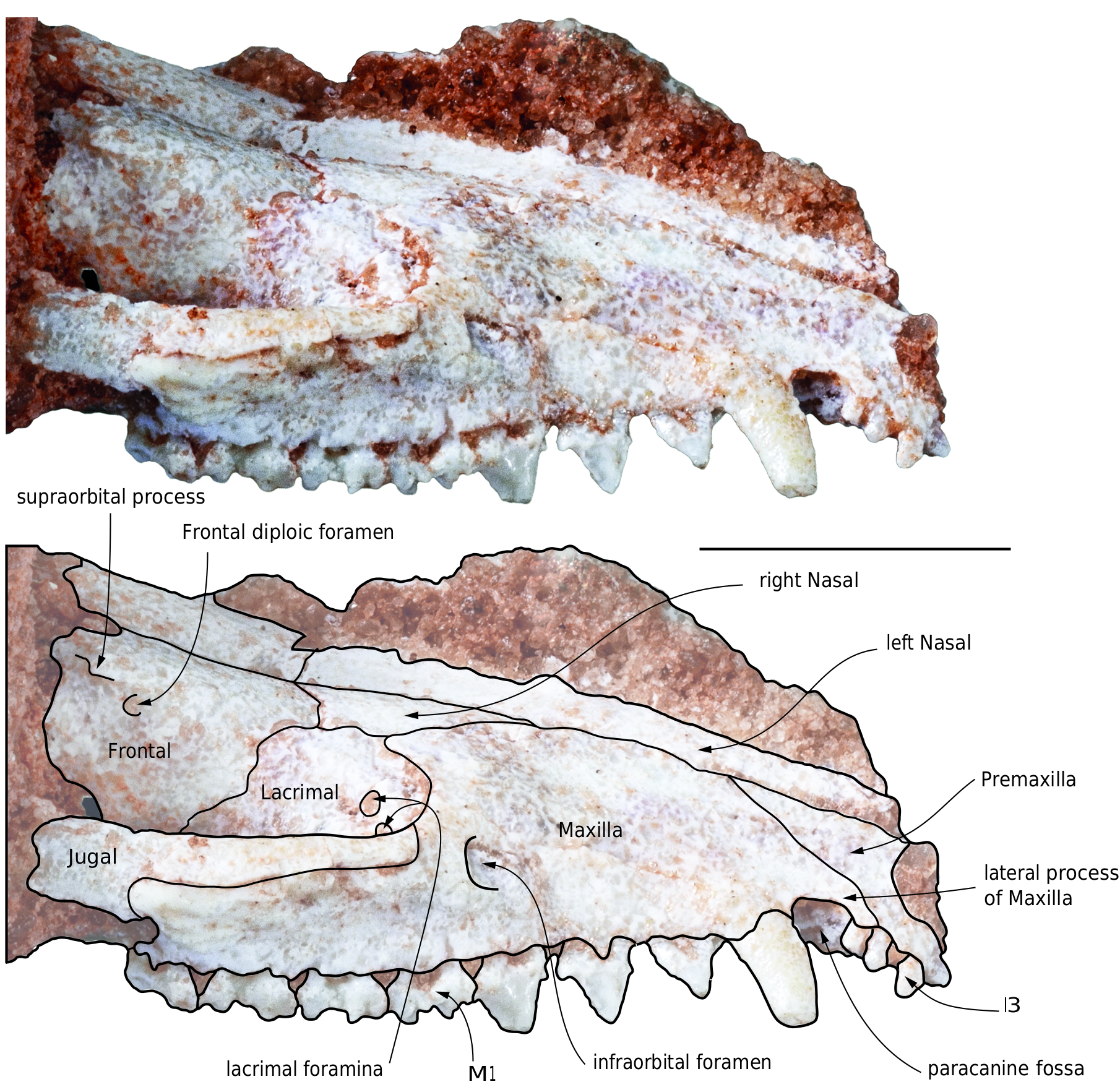
Lacrimal ( Figs 8 View FIG ; 9 View FIG ). The lacrimal is relatively large. It forms the anterior edge of the orbit, although it is mainly internal to it. The facial process is crescent-like and narrow. It does not strongly extend on the face as it does in the basal metatherians, deltatheroidans and sparassodonts (including Mayulestes ). It resembles the condition in Pucadelphys and extant didelphids. The internal portion of the lacrimal is three to four times larger than the facial process. As discussed above, the lacrimal has a long suture with the maxilla anteriorly, a small suture with the nasal dorsally with the frontal posterodorsally, with the palatine posteroventrally and ventrally, and a small one with the jugal ventrolaterally. On the anterior edge of the orbit, but internal to it, two lacrimal foramina open posteriorly. They are subequal in size and roughly circular to oval-shaped. They are positioned one almost lateral to the other along a subhorizontal (dorsomedial-ventrolateral) axis; in other words, the lateral lacrimal foramen is slightly ventrolateral to the medial one. No lacrimal tubercle is observed. A lacrimal tubercle is absent in Pucadelphys and Mayulestes . It is present Andinodelphys and variably present in some extant didelphids (e.g., Caluromys , Thylamys – personal observations). Within the orbit, the lacrimal forms the dorsal edge of the maxillary foramen.
Nasal ( Figs 8 View FIG ; 9 View FIG ). The nasals are elongated narrow bones, with sub-parallel edges in their anterior two thirds and which strongly widen in their posterior third. The anterior apices of the nasals are probably missing and it is not possible to observe if their anterior ends overhang the narial fossa or not. The nasal-maxilla suture is anteroposteriorly oriented up to the level of P3. There, it turns laterally at approximately 45° and turns posteriorly just before contacting the lacrimal. Posterior to this bone, the nasal contacts the frontal and the nasal-frontal suture turns medially at approximately 45°. At the medial third of the frontal width, the suture turns anteromedially at 90° and meets the sagittal plane. The posterior suture of the nasals, with the frontals, is distinctly W-shaped as in Pucadelphys and in most didelphids (e.g., Hyladelphys , Marmosa , Monodelphis , Metachirus , Didelphis , Lutreolina, Cryptonanus , Lestodelphys , Thylamys ). It differs from that of Andinodelphys in which it is markedly convex posteriorly. This posterior flared portion of the nasals is longer than wide, but not as elongated as in extant didelphids. It differs from the diamond-shaped morphology observed in Andinodelphys .
In its lateralmost region, the nasal contacts the lacrimal as is observed in sparassodonts and deltatheroidans. Posteriorly, the nasals extend approximately as far as mid-length of the orbit and remain well anterior to the supraorbital processes.
Jugal ( Figs 8 View FIG ; 9 View FIG ). The jugals are preserved only in their anterior part, which articulates with the maxilla and which borders the orbit ventrolaterally. On its lateral aspect, in didelphids, this region of the jugal bears a deep elongated fossa for the origin of the zygomaticus and levator labii superioris. No fossa is observed in this region of the jugal of Incadelphys . The levator labii superioris probably originated in the fossa in the maxilla below the maxilla-jugal suture, as discussed above, and the zygomaticus probably originated on the jugal as in Didelphis , although no fossa for this muscle is observed on the jugal of Incadelphys . The condition of the latter is similar to those of Andinodelphys and Pucadelphys and therefore clearly differs from that in extant didelphids.
Frontal ( Figs 8 View FIG ; 9 View FIG ). Dorsally, a small part of the frontals is preserved, which forms the interorbital bridge. Anteriorly, the frontals feature conspicuous anterior processes which wedge between the nasals anteriorly. Laterally, the frontal bears a small and sharp supraorbital process as in Andinodelphys and Pucadelphys . Anteroventral to the process is a small anteriorly-opening frontal diploic foramen ( Figs 8 View FIG ; 9 View FIG ). This foramen likely conveyed the frontal diploic vein, an emissary of the dorsal cerebral vein/dorsal sagittal sinus or a vein issued from the frontal diploe ( Thewissen 1989; Evans & de Lahunta 2013; Wible et al. 2004). This foramen is called supraorbital foramen by Novacek (1986) and Marshall & Muizon (1995), the frontal foramen by Evans & de Lahunta (2013), the foramen for the frontal diploic vein by Wible & Rougier (2000), Wible (2003), Wible et al. (2009) and Wible (2011), and the frontal diploic foramen by Thewissen (1989). In this paper, we retain the terminology “frontal diploic foramen”. Such a foramen is present in Pucadelphys , Andinodelphys , and generally in Recent didelphids.
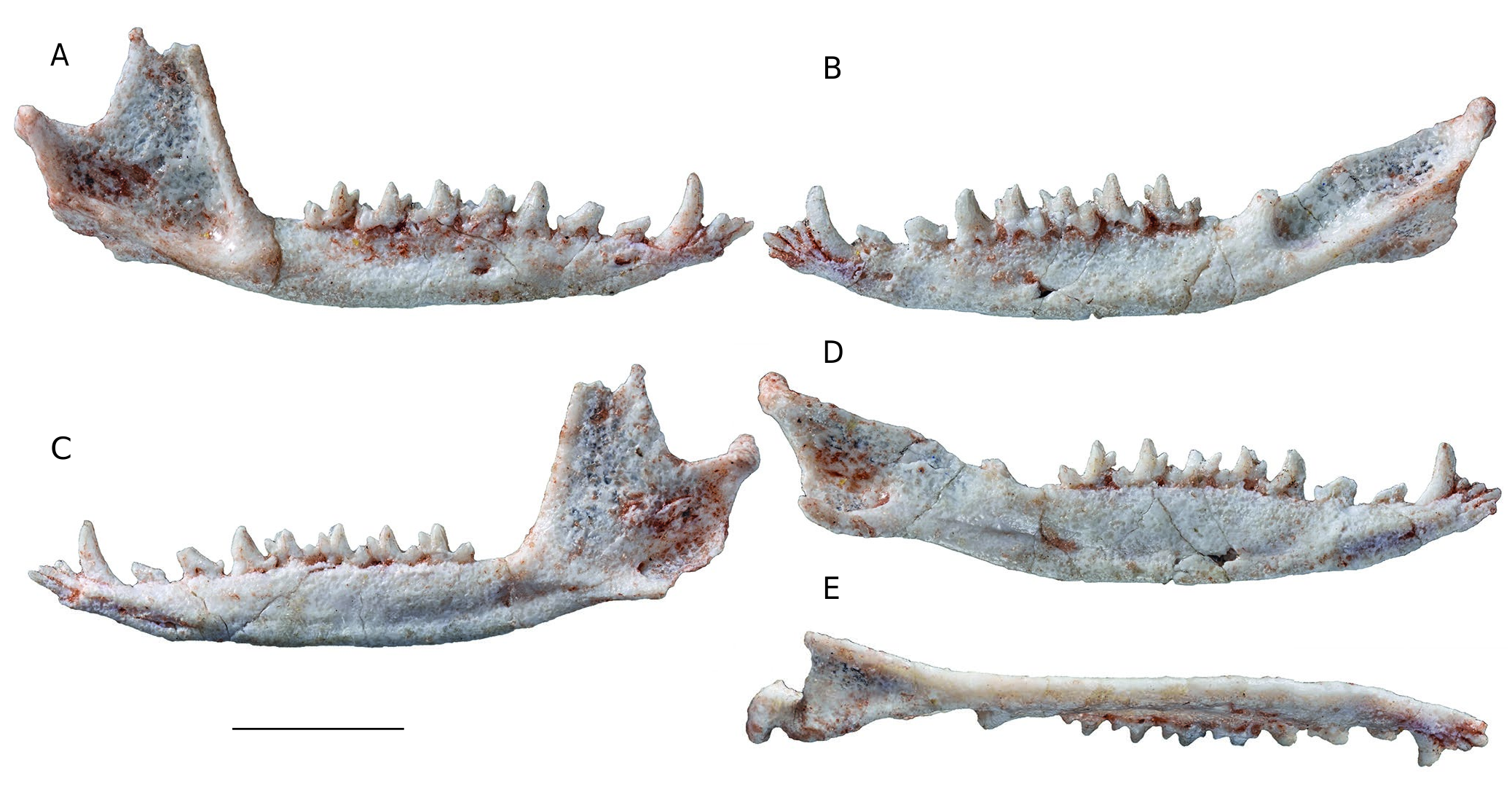
Dentary ( Fig. 10 View FIG ). The dentary of Incadelphys is very slender and gracile to a greater extent than all the Tiupampa metatherians for which a complete dentary is known ( Table 4 View TABLE ). The dentary of Incadelphys is proportionally even longer and lower than in Andinodelphys , which has a notably elongated snout ( Muizon & Ladevèze 2020). As seen in Table 4 View TABLE , the dentaries of Pucadelphys and Mizquedelphys are consistently shorter proportionally than those of Incadelphys and Andinodelphys and the stouter metatherian dentaries known at Tiupampa are those of Mayulestes and Allqokirus . Therefore, a gradient of rostrum slenderness is observed in the Tiupampa metatherians increasing from the slenderest morphology in Incadelphys to the stouter in Allqokirus-Mayulestes: Incadelphys < Andinodelphys < Pucadelphys < Mizquedelphys <Allqokirus-Mayulestes.
The corpus mandibularis slightly increases in depth posteriorly and has a maximum elevation below m2-m3. Then it slightly decreases posteriorly as far as the retromolar space. The ventral edge of the corpus is weakly convex, the convexity being maximum below m4. It is less convex than in the extant didelphids Didelphis and Caluromys , in which the corpus is generally deeper, with the deepest point located more posteriorly, below m3-m4. The condition of Incadelphys more closely resembles, in this respect, that of Andinodelphys . Three mental foramina are observed, a small one below i3 and, on the lateral aspect of the corpus, two large mental foramina are present, the anterior one below p1 and the posterior one below m1. The foramen observed in Andinodelphys between the dorsal edge of the symphysis and the lingual alveolar border of the canine is absent in Incadelphys . On the medial aspect of the corpus, the mandibular symphysis is unfused. It is elongated and low and extends posteriorly as far as a point below the anterior root of p2. The long axis of the symphysis is at an angle of approximately 19° to the horizontal axis of the tooth row (the alveolar border), which resembles the condition in Andinodelphys ( Muizon & Ladevèze 2020) . As in this latter genus, the condition of Incadelphys is related to the anterior tapering of the rostrum and procumbency of the incisors. It contrasts with the shorter rostrum of Pucadelphys (c. 22.4°) and especially of Mayulestes (29°) and Allqokirus (30°) (see Muizon & Ladevèze 2020: table 7). Among extant didelphids, the condition of Incadelphys resembles that of Metachirus , but differs from Didelphis and Caluromys , in which the symphysis is slightly less slanted.
A conspicuous mylohyoid groove is present and extends from a point below p3 as far as a point below the anterior end of the coronoid crest. Between the last molar and the base of the coronoid crest is a small retromolar space, which is slightly shorter than m4.
On the ramus, the coronoid process is large but proportionally smaller than in Didelphis , shorter (proximodistally) than in Caluromys , and approaches the size observed in Metachirus . Its apex is incomplete but little of the process is missing. Because its posterior edge is straight it is probable that its apex was not (or little) recurved posteriorly, as is observed in Andinodelphys and Pucadelphys . The straight posterior border of the coronoid process differs from the condition generally observed in didelphids, but resembles the straight posterior border of the coronoid process in Dasyurus . The coronoid crest, which forms the anterior edge of the process, is straight and proportionally thinner than the condition observed in Andinodelphys , in which it is salient towards its base and extends on the lateral aspect of the body. The condition of Incadelphys resembles that of Pucadelphys in this respect. The masseteric fossa is remarkably deep, especially posteroventrally, in the region anteroventral to the condyloid process, and anteroventrally, posterior to the coronoid crest. In the posteroventral region of the masseteric fossa, a sub-horizontal posterior shelf fossa projects laterally, being distinctly convex. The maximum lateral extension of this shelf is just ventral to the anteriormost point of the anterior edge of the condylar process, being more posterior than in Andinodelphys . The medial surface of the coronoid process is smooth and flat to slightly convex medially and bears no anterior crest.
Ventrally, the posterior crest of the coronoid process turns posteriorly and joins the articular condyle. This crest descends more ventrally than the condyle before reaching it and form a distinct notch between the two processes ( Fig. 10 View FIG ). It reaches the condylar process in its medial third and is approximately posterior to the m 4 in the axis of the tooth row. Therefore, the lateral two thirds of the condyle are lateral to the tooth row and overhang the posteroventral region of the masseteric fossa. The condition of Incadelphys is similar to that observed in Pucadelphys and Andinodelphys . Only one third of the transverse length of the condyle is medial to the tooth row. The condyle is strongly elongated transversely and cylindrical. Its articular surface is posterodorsally oriented. In its medial portion the condyle tappers being anteroposteriorly shorter. The lateral part of the condyle is buttressed by the ascending posterior end of the posterior shelf of the masseteric fossa.
On the posteromedial edge of the condylar process, just ventral to the point of junction between the coronoid and condylar processes, a robust ridge descends towards the posteromedial edge of the angular process. In lateral view, the articular surface of the condyle is located above the apex of the protoconid of m4, at a distance of approximately twice the height of the trigonid of m4. Therefore, the condyloid process is in a much higher (c. four times more elevated) position than in Andinodelphys and Pucadelphys , in which the distance is approximately the height of the talonid of m4.
The angular process is shelf-like and inflected medially, as in most metatherians ( Fig. 11E View FIG ). It is triangular in ventral view and approximately twice as long as wide posteriorly. Its posterior edge is slightly concave, and its medial angle forms a short but pointed triangular process more pronounced than in Pucadelphys and Andinodelphys but clearly differing from the sharp, posteriorly-projecting spur-like process observed in didelphids and, to a greater extent, in dasyurids. In Didelphis , the dorsal surface of the angular process receives the insertion of the internal pterygoid and its ventral aspect supports the superficial masseter ( Hiiemae & Jenkins 1969). Sanchez-Villagra & Smith (1997) have established categories of the diversity of the angular process in marsupials based on the ratio of “angular process length to angular process shelf length”. The ratio value calculated for Incadelphys is c. 0.83, which places it in the “shelf-like” category (Ratio> 0.81). However, it is noteworthy that this ratio is significantly lower than in Andinodelphys in which the ratio is 0.89. Therefore, the ratio calculated for Incadelphys reflects the length of the posteromedial process of the angular process, conspicuously longer than in Andinodelphys . Slightly posterior to the point of departure of the angular process, on its lateral side, is a small mandibular foramen. It is circular to oval-shaped and widens posteriorly. It is located ventral to the middle of the apex of the coronoid process.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
InfraClass |
Metatheria |
|
SuperOrder |
Pucadelphyda |
|
Order |
|
|
SuperFamily |
Pucadelphyoidea |
|
Family |
|
|
Genus |