
Isophya modestior Brunner
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3658.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:C02D1C74-25C0-41DD-B098-62098EB7B62A |
|
DOI |
https://doi.org/10.5281/zenodo.5617379 |
|
persistent identifier |
https://treatment.plazi.org/id/F26F3128-392F-FFB5-B1B0-0A94FB1C9EA1 |
|
treatment provided by |
Plazi |
|
scientific name |
Isophya modestior Brunner |
| status |
|
Isophya modestior Brunner von Wattenwyl, 1882
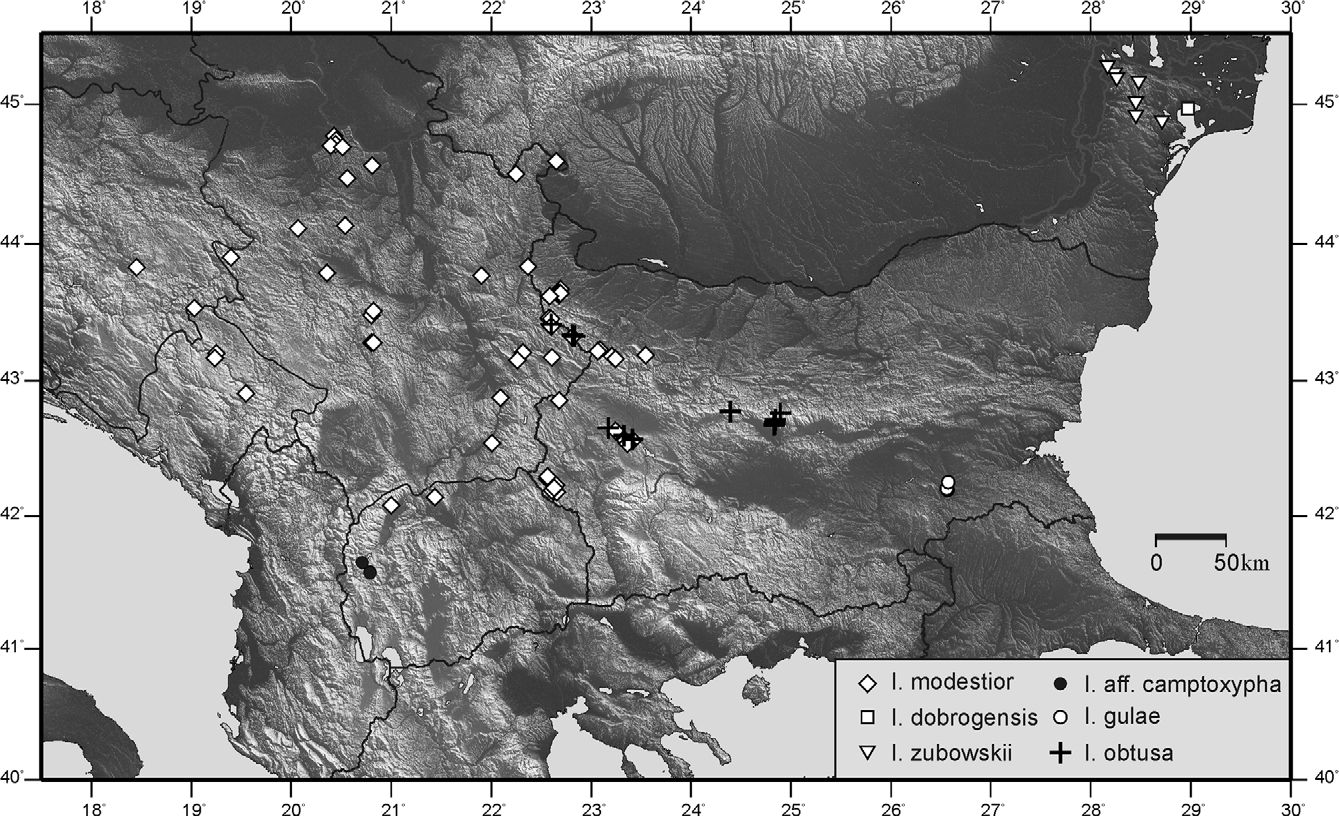
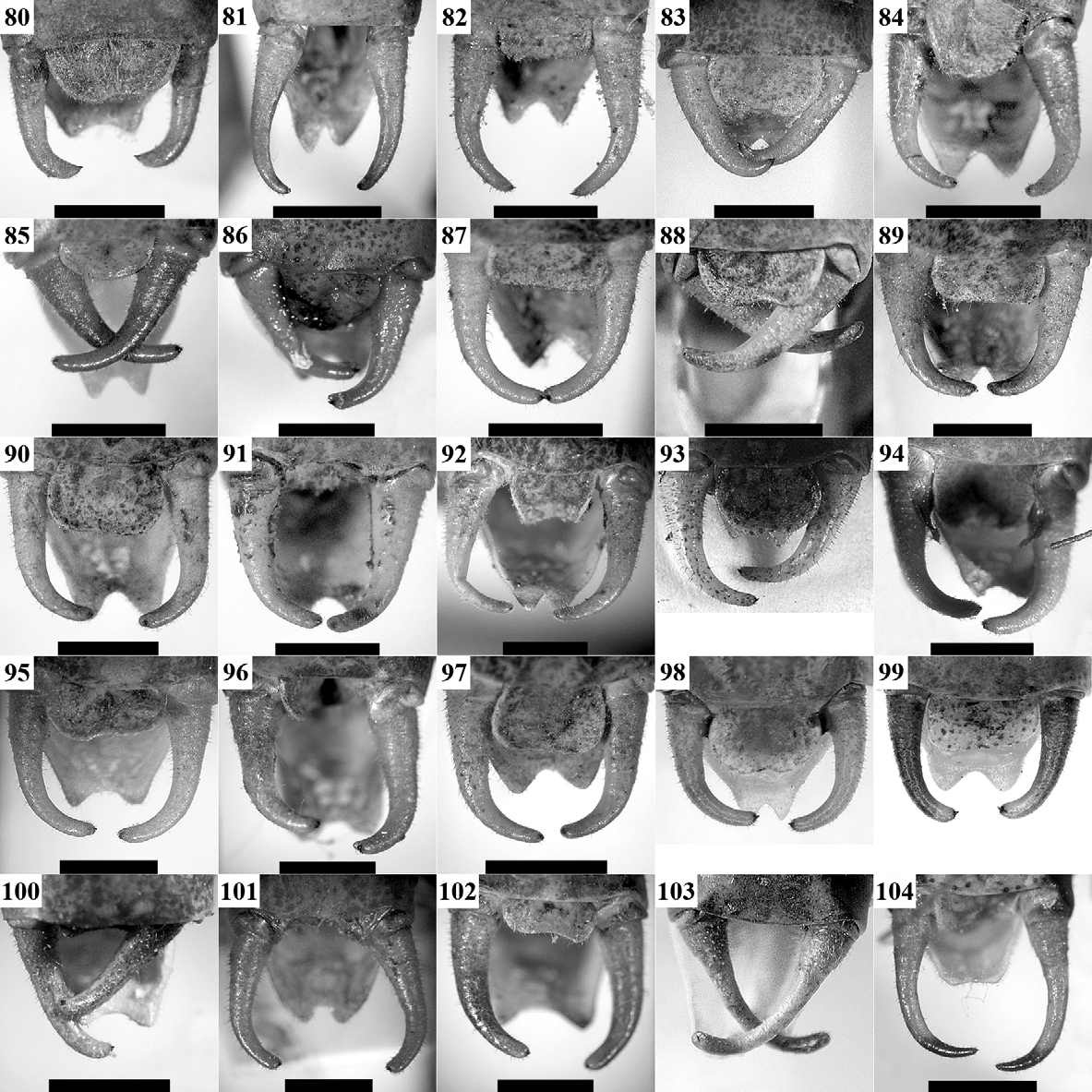
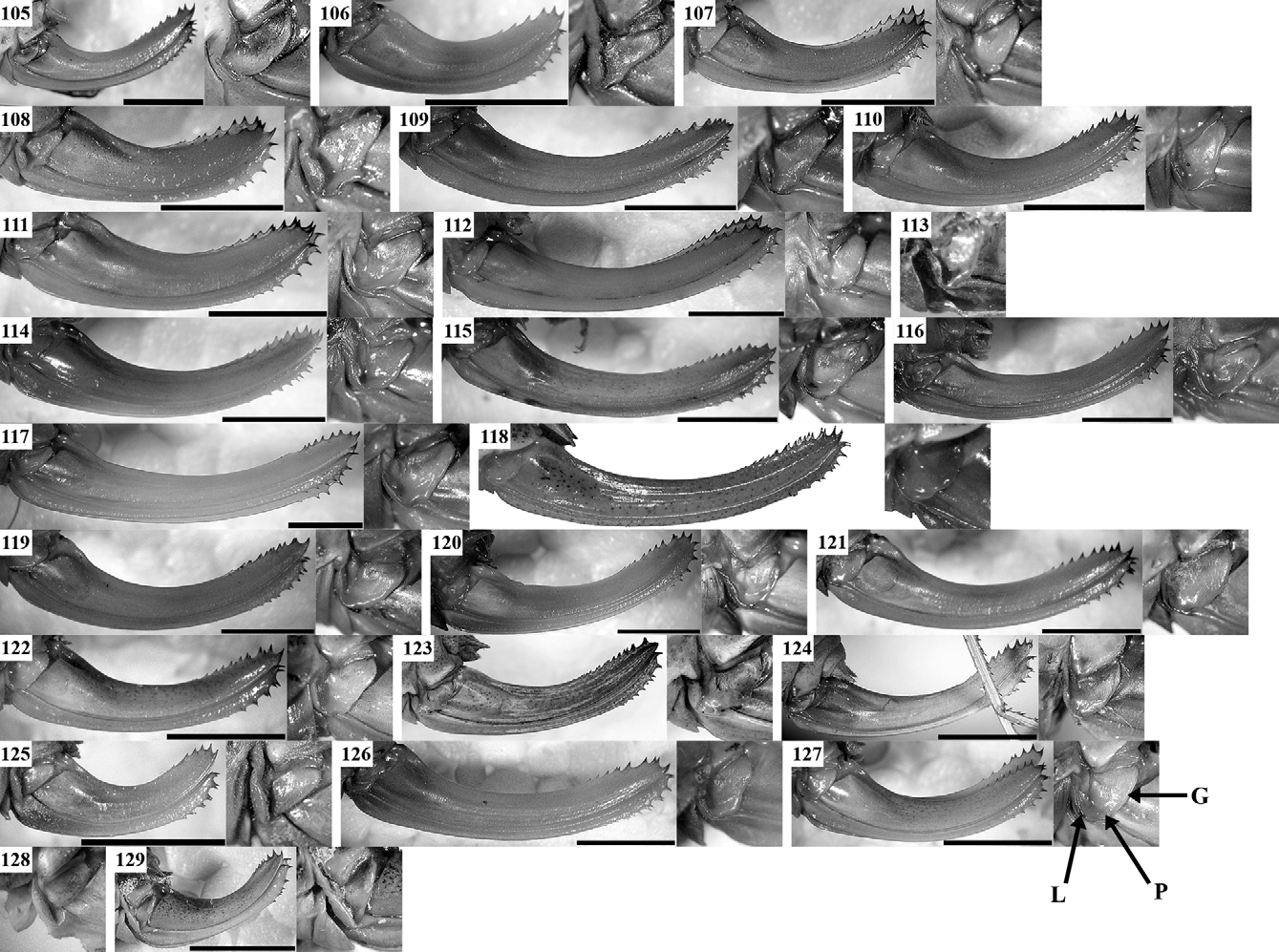
( Figs 19 View FIGURES 19 – 30 , 48, 72 View FIGURES 56 – 79 , 97 View FIGURES 80 – 104 , 122 View FIGURES 105 – 129 , 182, 183 View FIGURES 182 – 189 , 194 View FIGURE 194 )
Isophya modestior Brunner von Wattenwyl: Brunner von Wattenwyl 1882 (sp.n.). Isophya fusconotata Brunner von Wattenwyl: Brunner von Wattenwyl 1882 (sp.n.).
Morphological description: Brunner von Wattenwyl 1882; Bey-Bienko 1954; Harz 1969; Heller et al. 2004; Orci et al. 2005. Karyotype: Warchałowska-Śliwa et al. 2008. Bioacoustics: Heller 1988; Fontana et al. 2002; Nagy et al. 2003; Heller et al. 2004; Orci et al. 2005; Ingrisch and Paviċeviċ 2010, 2012.
Variation: Few geographically delimited populations exhibit differences in morphology (number of stridulatory teeth, Heller et al. 2004), song (see below), the type of X-chromosome (acro- or subacrocentric) (Warchałowska-Śliwa et al. 2008), and sequences of some genes (own unpublished information).
Bioacoustics: Three populations have been studied bioacoustically.
The Bulgarian population from Vitosha Mt. is characterised by a song ( Fig. 182 View FIGURES 182 – 189 ) of groups of two syllables separated by intervals of 2.5–10 s (mean 4.8±2.3; n=12). The first syllable in a group is usually shorter than the second one due to shorter average impulse period (about 0.4 ms) (the number of impulses is the same). At 26–27°С the syllables lasted 255–320 ms (mean 287±17; n=26) and included 88–97 impulses (mean 92±3; n=26). The impulse period lasted 2–7 ms (average 3 ms), being shortest around the middle of the first half of the syllable and longest at its end. The population from Western Stara Planina Mts showed a song (at 27.5°С) of many loosely following each other syllables separated by intervals of 10–20 s or more. The syllables showed similar characteristics as that described above.
The Serbian population from the region of Novi Sad (Fruska Gora, 280 m, 45.1870º N, 19.8130º E; outside the borders of the Balkan Peninsula but shown here for comparison) showed a song ( Fig. 183 View FIGURES 182 – 189 ) of groups of few syllables (usually 3–5) separated by intervals of 2–5 s. The syllable consisted of main part of 150–250 ms and afterclicks; its total duration is 280–350 ms. The impulses in the main part are denser than in the Bulgarian population.
Karyology (Warchałowska-Śliwa et al. 2008 and unpublished data): the Bulgarian population (specimens studied from the surroundings of Tran) showed 2A type of X-chromosome with telomeric C-bands (heterochromatin) on both arms; the Serbian population differed in possessing 1A type of X-chromosome.
Morphology: All studied populations did not exhibit morphological differences. The stridulatory file had respectively 182 (W Stara Planina Mts), 200 (Vitosha Mt.), and 211 (Novi Sad) stridulatory teeth, which coincides with the data known for the southern range of this species (see e.g. Heller et al. 2004).
Distribution ( Fig. 194 View FIGURE 194 ) and phenology: The species is known from the Northwestern Balkan Peninsula and the neighbouring parts of Central Europe (SW Romania, NW Bulgaria, N Macedonia, Montenegro, Bosnia and Herzegovina (new for this country), Serbia, Croatia, Slovenia, NE Italy, Austria, Hungary and possibly Slovakia after Heller et al. 2004; Szövényi and Puskás 2012). The species inhabits mesophyte scrub and grass vegetation to open meadows (at the upper limit of distribution) between 100 (in the northern part of its range) or 500 (in Bulgaria) to 1800–1900 m alt. (Ossogovska Planina Mt. in Bulgaria). Nymphs—IV–VI(–VII), imago—VI–VIII.
Notes to the literature distribution data: Buresch and Peshev (1958) reported I. modestior from Assenovgrad and Chepelare. It is still not clear which species may be concerned in this case but most probably this is I. rhodopensis . The localities in Rila (Parangalitsa Reserve, 18.06.1984, 1 Ƥ, HMB) and Pirin (Pirin Peak, 20.08.1986, 1 Ƥ, HMB) as labelled by Andreeva are doubtful and may concern mistake. Thus, they are not mapped or included in the list of material. The data by Peshev (1974b) for Vratsa (23.06.1964) and Stakevtsi Village (12.08.1966) are referred to I. miksici on the basis of the revised material (see Appendix). Köhler (1988) recorded I. modestior from Northwestern Pirin Mts but this record may concern one of the species that occur in this region and may be misidentified with I. modestior : I. andreevae , I. rhodopensis leonorae or I. bureschi .



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |