
Jamaha playa Bahder & Bartlett, 2023
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5263.2.5 |
|
publication LSID |
lsid:zoobank.org:pub:4900801D-0D22-4FD6-9ABF-EFC912272CCA |
|
DOI |
https://doi.org/10.5281/zenodo.7801062 |
|
persistent identifier |
https://treatment.plazi.org/id/BD4987A7-FF96-FF80-FF11-3A1FFD4F84F5 |
|
treatment provided by |
Plazi |
|
scientific name |
Jamaha playa Bahder & Bartlett |
| status |
sp. nov. |
Jamaha playa Bahder & Bartlett sp. nov.
( Figures 2 View FIGURE 2 –9)
Type locality. Snow Hill , Portland Parish, Jamaica (18.197197, -76.525658) ( Fig. 1 View FIGURE 1 ) GoogleMaps .
Etymology. The specific epithet ‘ playa ’ is the Spanish word for beach, based on the habitat where the specimen was collected. The specific name is intended to be indeclinable.
Diagnosis. Vertex (dorsal view) parallel-sided and medially concave, apex weakly convex, posterior margin deeply concave. Frons elongate, bearing median carina, roughly rhomboid, widest dorsad of frontoclypeal suture. Forewings with Sc+RA, MP and CuA all originating near apex of basal cell. Hind tibiae with a lateral spine. Each lateral margin of pygofer opening bearing 2 large posterior processes, a large dorsal spatulate process and an elongate ventral process. Aedeagus bearing multiply branched subapical lateral processes (left side with 4 apices, right side with 3). Anal tube in lateral view with apex downcurved and broadly expanded into large truncate apices.
Description. Elongate (6.94 mm with wings, Table 3 View TABLE 3 ), entire body stramineous with fuscous wash over entire body, abdominal tergites orange/rust colored, wings generally fuscous with costal region transparent and anal region mottled, fuscous patch near apex on posterior margin.
Head. In dorsal view, head (including eyes) just wider than pronotum. Vertex quadrate, wider than long at midline, lateral margins subparallel, raised (medially depressed), median carina evident, anterior margin convex, transverse carina at fastigium, posterior margin, strongly concave ( Fig. 3A View FIGURE 3 ). In lateral view, head weakly projected in front of eyes, dorsum roundly declinate to fastigium, angled at transverse carina, face weakly convex ( Fig. 3B View FIGURE 3 ). In frontal view, vertex concave, transverse carina at fastigium weakly concave, face (including clypeus) pentagonal, lateral margins of frons keeled, expanding from fastigium approximately to level of antennae, then constricted to frontoclypeal suture; median carina distinct to frontoclypeal suture. Frontoclypeal suture sinuate, medially with concave dimple, laterally convex to margin; clypeus triangular ( Fig. 3C View FIGURE 3 ). Compound eyes oval, ventrocaudal margin flattened (weakly concave near antenna), lateral ocelli distinct beneath eyes. Antennae below eyes and caudad of lateral ocelli, scape very short and ring-like, pedicel ovoid, bearing many sensory plaques, flagellum bristle-like with bulbous base.
Thorax. Pronotum in dorsal view widest at midline, strongly narrowing laterally, anterior and posterior margins keeled, anterior margin strongly convex, posterior margin strongly concave, median carina present, disc bearing 3 indistinct tubercles along posterior margin on each side of midline ( Fig. 3A View FIGURE 3 ); in lateral view, paradiscal region narrow, strongly sinuate on anterior and posterior margin anterior margin keeled ( Fig. 3B View FIGURE 3 ). Mesonotum tricarinate, median carina reaching scutellum, lateral carinae strongly diverging from median carina near anterior margin, then abruptly much weaker, angled parallel to median carina to hind margin ( Fig. 3A View FIGURE 3 ). Fore and mid tibia approximately 2/3 length of hind tibia, both lacking lateral spines, hind tibiae with single lateral spine and apical spinulation of tibiae and tarsal segments 8-8-2.
Forewing. Forewing thickened, elongate, leading margin sinuate, weakly convex to weak inflection at stigmatal region, costal margin straight from scutellar margin to apex of clavus ( Fig. 4 View FIGURE 4 ); wing apex rounded, slightly projected in region of MP 1+2. Vein reticulations throughout forewing (including clavus), strongest in distal third. Apex of clavus in distal ¾ of forewing, fusion of Pcu and A 1 in distal third of clavus, composite vein reaching wing margin at claval apex. Sc+RA, MP and CuA all originate near apex of basal cell. Postcostal cell with 7 faint costal crossveins to subcostal posterior region, then 7 stronger crossveins in subcostal posterior region. RP ending with 3 terminals. MP forked near wing midlength (forming cell C3), apical region bearing 12 apical veins (between last RP vein and postclaval margin). CuA bearing 3 terminals.
Male terminalia. Pygofer in lateral view irregular, narrowest dorsally ( Fig. 5 View FIGURE 5 ), posterior margin convex bearing two large processes; dorsal process broadly spatulate with rounded apex, taller than wide; ventral process (the genus-level diagnostic feature) elongate, narrow, apex bluntly pointed. Gonostyli broad, rounded at apex, exceeding both processes of pygofer, capitulum short and broad, lateral tooth a subapical ridge ( Fig. 5 View FIGURE 5 ). Aedeagus bilaterally asymmetrical, shaft simple, terete, weakly upcurved bearing multiply branched subapical lateral processes (dorsolateral phallobase lobes, ventral hooks absent); apical processes (A1 & A2) hooked cephalad, short; middorsal processes (A3 &A4) elongate, slender, curved dorsad, irregularly sinuate; basal processes on left lateral side apically bifurcated (A5 & A7) into robust processes similar in length to A1 and A2; basal process on right lateral side (A6) lacking ventral bifurcation ( Fig. 6 View FIGURE 6 ). Anal tube in lateral view narrow basally, subparallel on dorsal and ventral margins, downcurved ventrally past midpoint, constricting then expanded to very broad truncate apices ( Fig. 5 View FIGURE 5 ). Paraproct thin and elongate.
Female. Unknown.
Type material. Holotype, ♁: “ Jamaica, Portland Parish / nr. Snow Hill (restaurant) / 15. II.2022 / sweeping weeds on beach / Coll.: B.W.Bahder // Holotype / Jamaha playa ♁” ( FLREC).
Plant associations. Unknown, collected sweeping herbaceous weeds on the beach.
Distribution. Jamaica (Portland Parish), near Snow Hill.
Sequence data. For J. playa sp. nov., a 531 bp product was generated for the COI gene (GenBank Accession No. OQ120945) and a 1,402 bp product for the 18S gene (GenBank Accession No. OQ116784). Based on data available for other taxa within Nogodinidae for the COI and 18S gene, J. playa sp. nov. resolves adjacent to Goniospara spp. with weak bootstrap support (56) ( Fig. 7A View FIGURE 7 ). For the 18S gene, of the available taxa, J. playa sp. nov. resolved closest to the Colpoptera sp. (from this study) with strong bootstrap support (100). The pairwise comparison generated based on 18S data demonstrates that J. playa sp. nov. differs from Colpoptera sp. by 4.3% and on average, differs by 5.1% (±0.1) from other genera within the Nogodinidae .
Remarks. The elongate projection on the lateral margin of the pygofer opening places J. playa sp. nov. in the genus Jamaha . Features of the male terminalia seem to offer the clearest features for species recognition. The terminalia of J. elevans and J. chrysops were both illustrated (as Colpoptera ) in Fennah (1955). Jamaha chrysops lacks the dorsal spatulate process of the pygofer found in J. playa sp. nov. and differs in the form of the subapical lateral processes of the aedeagus and the form of the anal tube (viz. Fennah 1955, Figs 1n, 1o View FIGURE 1 ). Jamaha elevans is more similar to J. playa sp. nov. in that both possess a rounded lobe near the dorsal margin of the pygofer in lateral view, however, this lobe appears larger and constricted at the base in J. playa sp. nov. compared to J. elevans ( Fennah 1955, fig 2q). Also, J. elevans has fewer branches of the subapical lateral processes (2 apices on the right side, 3 on left, versus 3 on right, 4 on the left in J. playa sp. nov.). Distant (1910) considered C. rugosa and J. elevans synonyms, however, this was not supported in later works. We examined syntypes of Colpoptera rugosa from the California Academy of Sciences collection to determine whether our new species might be attributed to that taxon. While we were able to exclude C. rugosa , we discovered that the syntype series were of mixed identity, which invoked a cascade of issues that we felt were better contended with in a separate manuscript.
Based on the pairwise comparison, the level of variability between Jamaha and Colpoptera is consistent with intergenic levels of variability (based on 18S) based on the taxa available for analysis. Molecular resources for the group are limited so no definitive conclusions can be drawn from the analysis performed, however, these data were intended as preliminary, and as new species are discovered, new molecular data are generated, and previously described taxa are analyzed, a clearer assessment of the relationships of species and genera in the Nogonidinidae will be possible.
Some of the values presented as intergeneric differences are low and likely more indicative of intrageneric levels of variability. It is unclear if this is due to errors in the taxonomy of the groups or errors in the identification of the specimens uploaded to GenBank, or both.
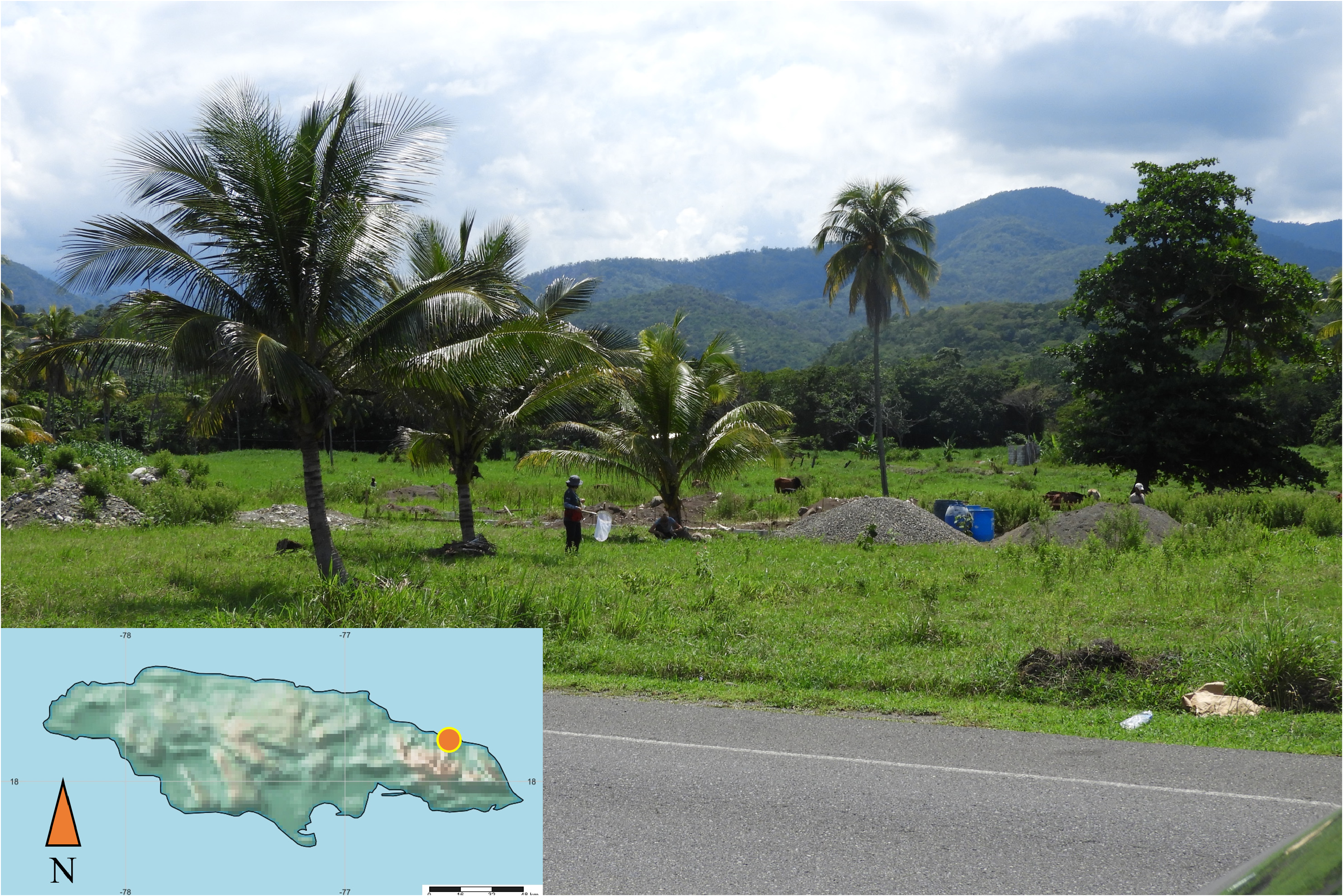

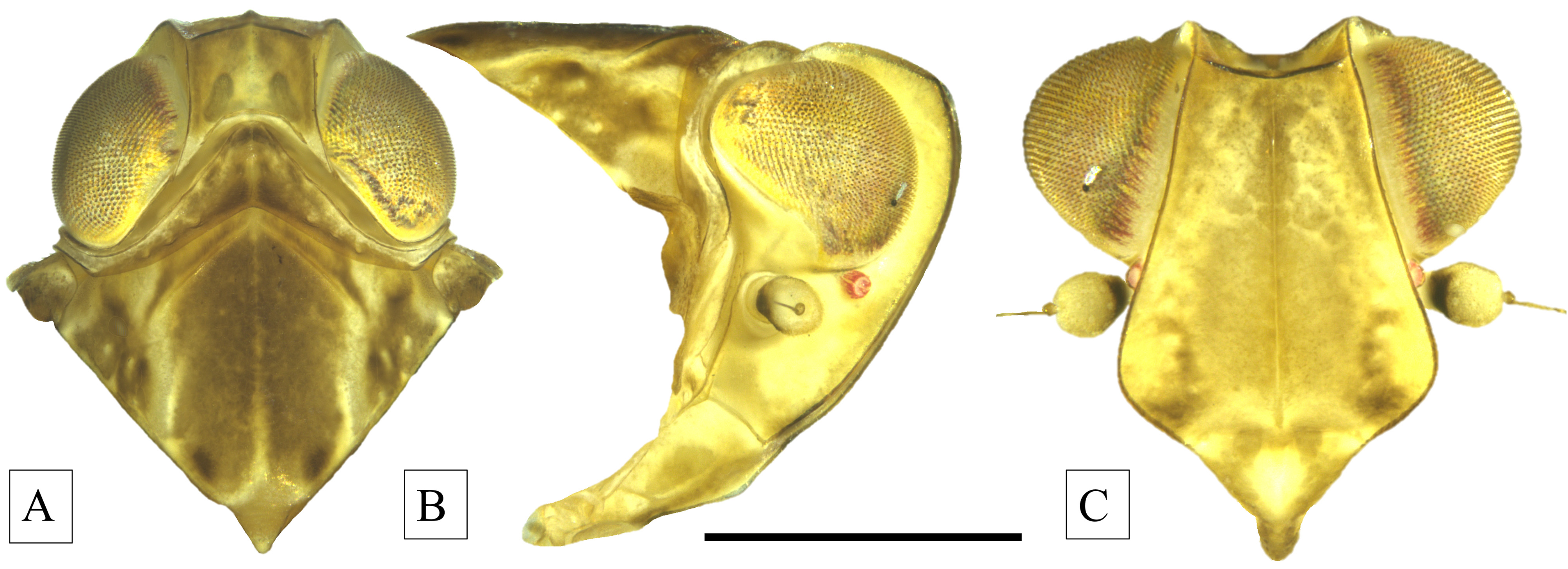
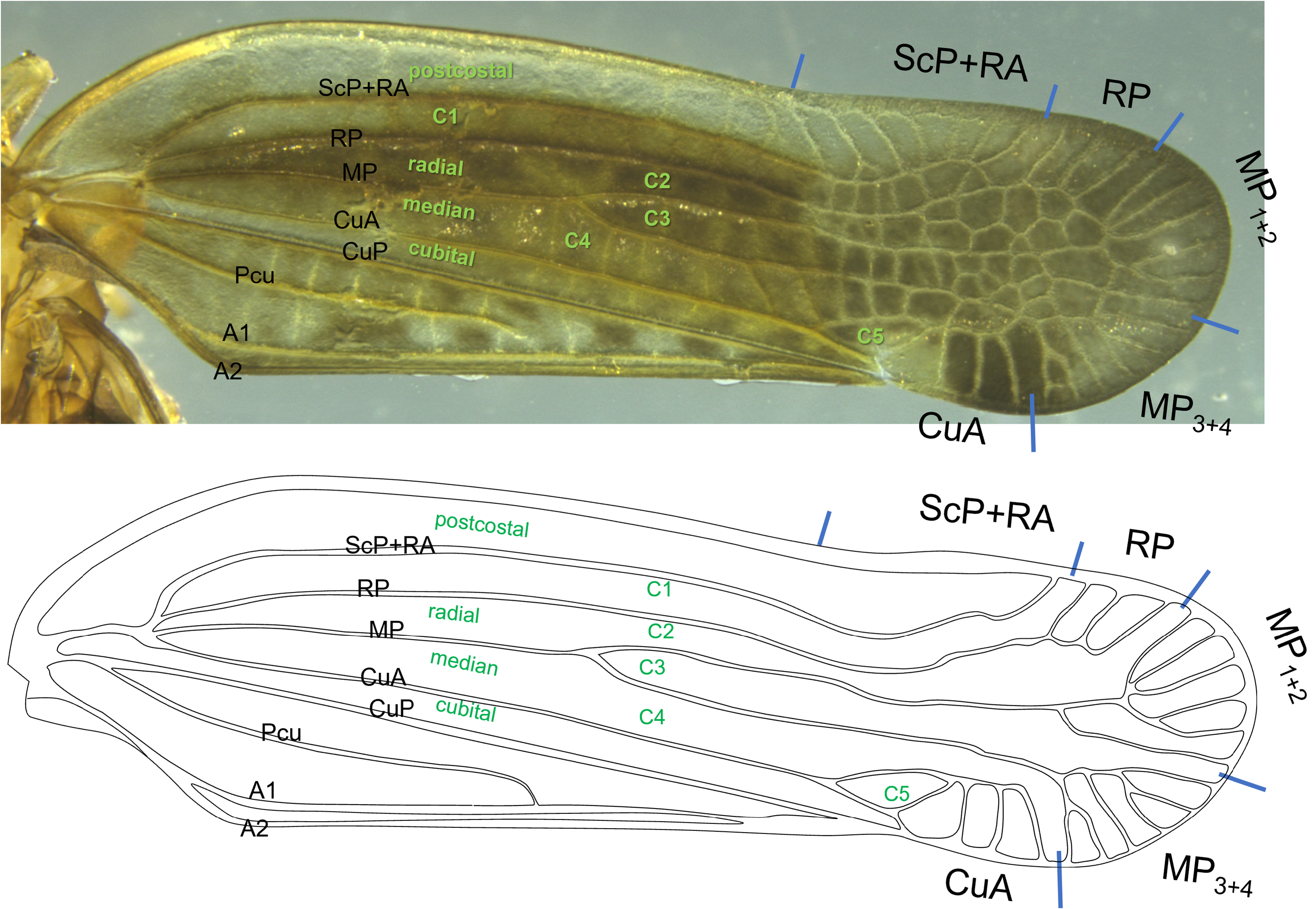
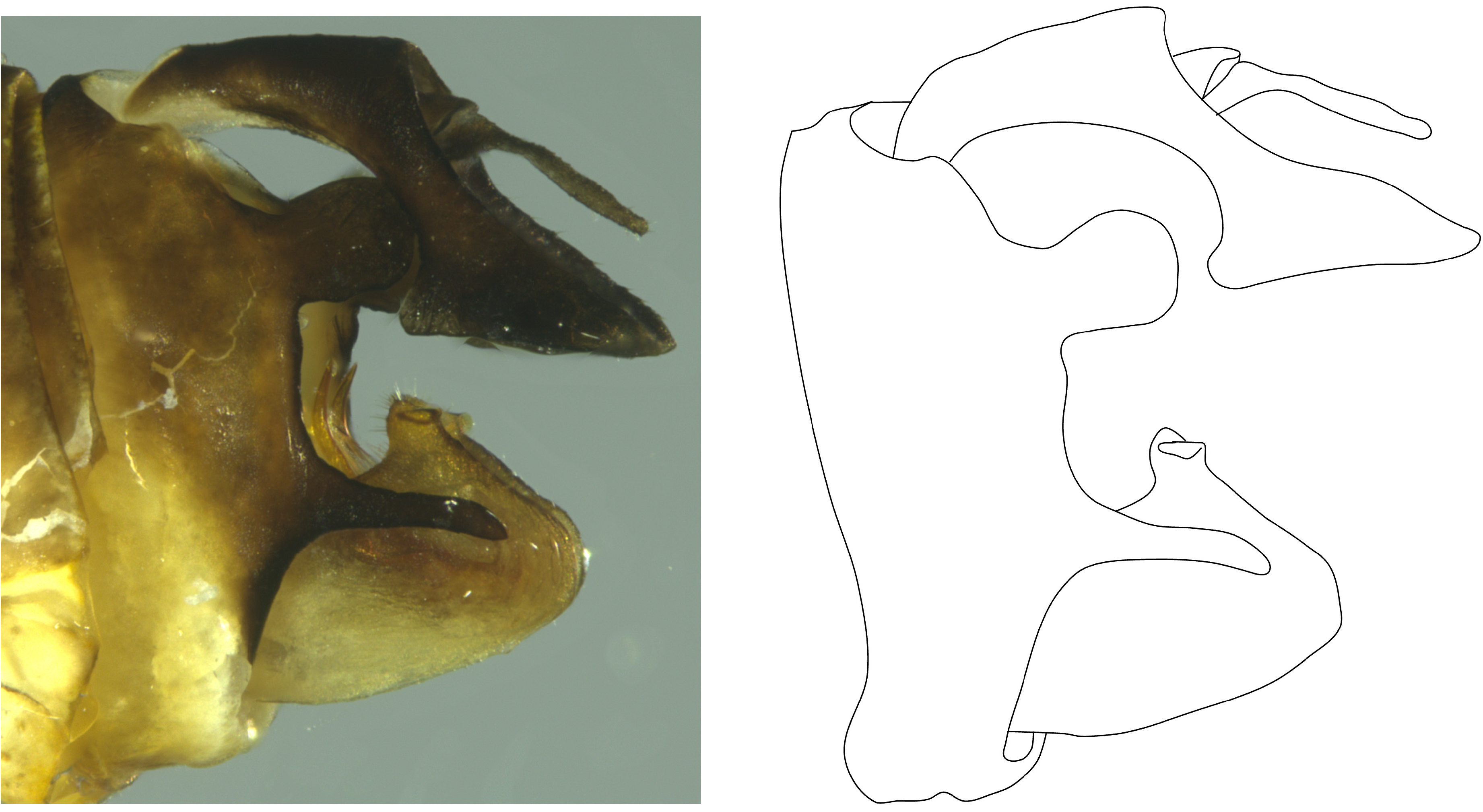
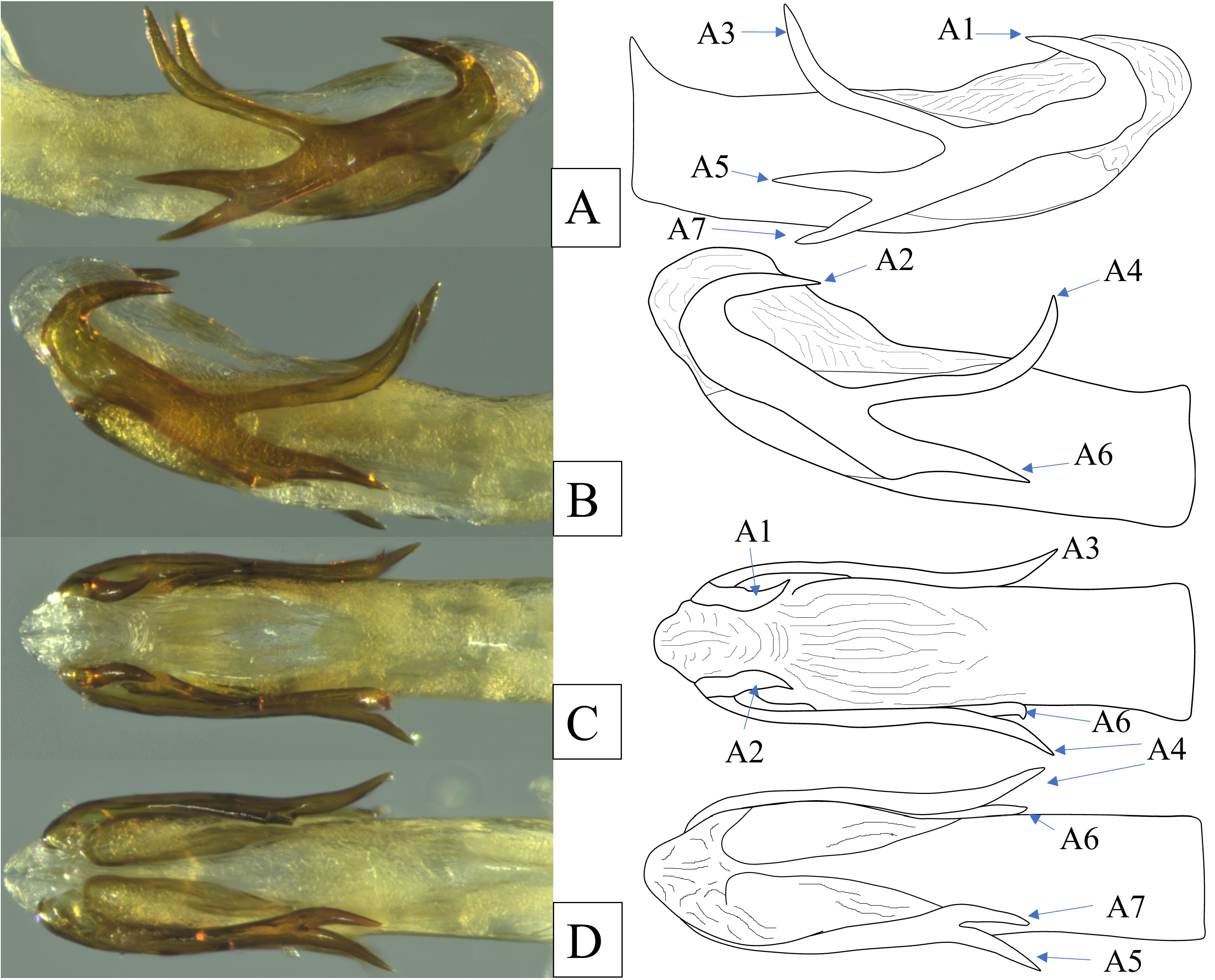
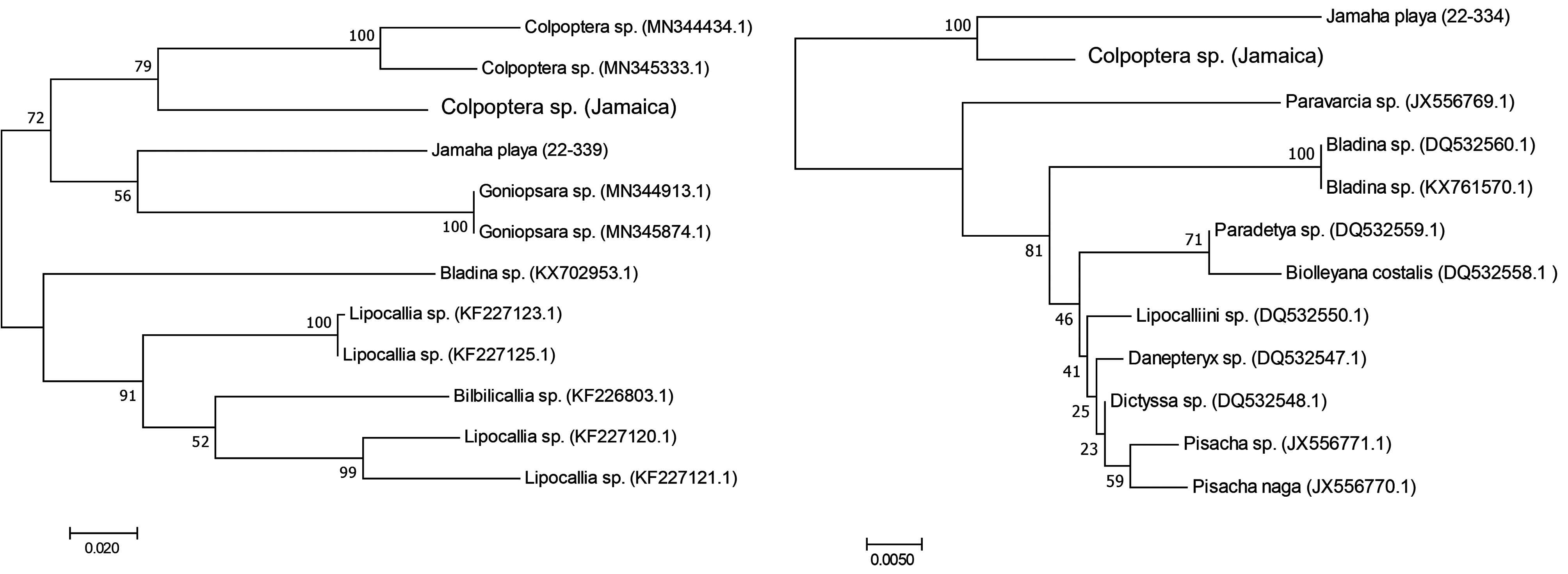
TABLE 3. Biometric data for Jamaha playa sp. nov.; measurements in mm. Character Measurement (n=1) Body length, with wings 6.94 Body length, no wings 4.33 Forewing length 5.98
| Character | Measurement (n =1) |
|---|---|
| Body length, with wings | 6.94 |
| Body length, no wings | 4.33 |
| Forewing length | 5.98 |
| Vertex length | 0.32 |
| Vertex width, basal margin | 0.53 |
| Vertex width, distal margin | 0.45 |
| Pronotum length, midline | 0.30 |
| Mesonotum length, midline | 1.21 |
| Mesonotum width | 1.38 |
| Frons width, dorsal margin | 0.45 |
| Frons width, clypeal suture | 0.51 |
| Frons width, widest | 0.92 |
| Frons width, narrowest | 0.45 |
| Frons length, midline | 1.02 |
| Clypeus length | 0.75 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SuperFamily |
Fulgoroidea |
|
Family |
|
|
SubFamily |
Colpopterinae |
|
Tribe |
Colpopterini |
|
Genus |