
Jodis orientalis Wehrli
|
publication ID |
https://doi.org/ 10.5281/zenodo.178403 |
|
DOI |
https://doi.org/10.5281/zenodo.5662166 |
|
persistent identifier |
https://treatment.plazi.org/id/BE05C364-B301-F828-76F2-F8C25E2919BD |
|
treatment provided by |
Plazi |
|
scientific name |
Jodis orientalis Wehrli |
| status |
|
Jodis orientalis Wehrli , stat. n.
( Figs 1, 2 View FIGURES 1 – 5 )
Jodis putata View in CoL f. orientalis Wehrli, 1923 , Iris 37: 62. Syntypes: 1 ɗ, 4 Ψ (ZFMK), "Mokanschan, Schanghai" [ China: Zhejiang, Mogan Shan Mnt., 50 km SW Hangzou]. The name is available based on ICZN (1999) article 45.6.4. because the context of the work does not clearly indicate that the name was proposed for an infrasubspecific entitity.
Iodis putata orientalis: Prout, 1935: 20 , pl. 6a (in part?).
Jodis putata orientalis: Parsons et al., 1999: 528 View in CoL .
Jodis angulata Inoue, 1961 View in CoL , Insecta Japonica 1(4): 53, text figs 116, 118, pl. 4, figs 91, 92, syn. n. Holotype, ɗ (BMNH), Japan: Funakoshi, Yokosuka.
Jodis angulata: Inoue, 1977: 234 View in CoL ; Inoue, 1982: 434, pl. 59: 2, Cat. 2201; Oh & Park, 1988: 165, fig. 23; Shin, 1996: 9; Kim et al., 2001: 30, fig. 24, pl. 2, fig. 6 (as " Jodis putata View in CoL ", misprint).
Material. Syntype, male, labelled: “Mokanshan / b. Hang-tschou”; “ orientalis / Wehrli / Jodis putata / ɗ Holo Type ” ( ZFMK).
Taxomomy. The examined syntype (originally labelled as “ Holotype ”) of Jodis putata f. orientalis differs from true Jodis putata (Linnaeus, 1758) in facies and male genitalia. Based on these features, it is undoubtedly a separate but congeneric species. It differs from J. putata by the more regular dentition of the postmedial transverse lines, which are not incurved markedly between veins M1 and M3 on the forewing. The male genitalia of Jodis putata f. orientalis differ by possessing a sclerotized dorsal and medial processes (harpe) on the valva and by the different shape of the eighth sternite, which forms two long posterior processes lacking scobination. The shape of the uncus, socii, gnathos, and transtilla, the outline of the valve, and the structure of aedeagus are all typical for Jodis .
The syntype conforms well to the original description, the figure of the male genitalia, and the images of the adult of Jodis angulata provided by Inoue (1961). The harpe on the original drawing of the male genitalia ( Inoue, 1961: fig. 116) is somewhat narrower and more rounded at the outer margin in comparison with that of the syntype of Jodis putata f. orientalis ( Figs 3–5 View FIGURES 1 – 5 ). However, this may be the result of slide mounting: the harpe in its natural position is trough-shaped but “unrolled” in the slide preparation. Therefore, Jodis angulata Inoue, 1961 , syn. n., should be treated as a junior synonym of Jodis orientalis Wehrli, 1923 , stat. n.
Based on the new synonymy, previous records of Jodis putata f. orientalis as an East Asian subspecies of Jodis putata should be regarded as misidentifications (e.g., Inoue, 1946b: 44; Inoue, 1956: 234; Inoue, 1961: 52; Inoue, 1977: 234; Inoue, 1982: 433; Viidalepp, 1976: 846; Viidalepp, 1996: 63; Kim et al., 2001: 29; Hausmann, 2001: 177). The true status of East Asian populations of Jodis putata is unresolved, but following preliminary analyses, differences between European and Far Eastern specimens appear insufficient for recognition as separate subspecies.
Distribution. East China (Zhejiang), Korea (Central, South, Jeju), Japan (Honshu, Shikoku, Kyushu).
Aoshakuna Matsumura , stat. nov.
Aoshakuna Matsumura, 1925 View in CoL , J. Coll. Agric. Hokkaido Imp. Univ. 15: 156. Type-species: Aoshakuna sachalinensis Matsumura, 1925 View in CoL , by original designation.
Nipponogelasma Inoue, 1946 View in CoL , Bull. lepidopt. Soc. Japan 1: 1, syn. n. Type-species: Gelasma immunis Prout, 1930 View in CoL , by original designation.
Diagnosis. The antennae are bipectinate in the male and filiform in the female. In the forewing R1 originates from the discal cell, is very short, and is fused with Sc for a long distance, and M3 and Cu1 are stalked; in the hindwing Rs and M1 are stalked, M3 and Cu1 are stalked, and the margin is slightly angled at M3. The frenulum is present in the male and absent in the female. The male hindtibia has a short distal projection with an androconial tuft. The abdomen lacks crests.
Male genitalia. The socii are similar to the uncus in shape and length; the valva has a strong, scobinate, pointed, arched dorsal process; the transtilla is an entire, narrow, arched bar between the valvae; the aedeagus has a long, posteriorly-directed, scooplike projection near the middle; and the eighth abdominal sternite has a needlelike process on the posterior margin.
Female genitalia. The lamella antevaginalis is large and subtriangular; the lamella postvaginalis is Wshaped, with the lateral arms lobelike, dilated posteriorly, and with the median angle tonguelike and deflected ventrally; and the corpus bursae is elongate, simple, with a large sacklike posterior dilation lateroventrad of the ductus bursae.
Taxonomy. The taxonomic status of the generic name Aoshakuna was uncertain because the type species, Aoshakuna sachalinensis , was described from a single female without a genitalia dissection. Prout (1934) nominally synonymized Aoshakuna with Chlorissa Stephens, 1831 , by including Aoshakuna sachalinensis in Chlorissa , but he also indicated that it “may well be a Chlorissa , possibly however a small Gelasma ” ( Prout, 1934: 15). While reviewing Matsumura’s collection, Sato (2001) found the holotype of Aoshakuna sachalinensis with a missing abdomen and the wing colour faded to yellow, with the lines almost gone. He compared it with Japanese Chlorissa species but refrained from associating it with any of them, noting that “material ... from Sakhalin is needed for further studies”; he retained Aoshakuna as a synonym of Chlorissa . In addition to the original description, Sato mentioned that the third segment of labial palpi is as long as in Chlorissa obliterata (Walker, 1863) .
In analyzing the systematic position of Aoshakuna sachalinensis , I collected key characters of the taxon from Matsumura’s original description and his original black-and-white photograph of the holotype, which was taken from the specimen before it faded, and addition characters provided by Sato. These characters are as follow:
1) wingspan 24 mm (the original description);
2) wing colour olive-green, fringe whitish (the original description);
3) forewing with transverse lines white; border with green deeper than ground colour; slightly wavy, postmedial line curved outward at Cu1 (originally “vein 3”) and curved inward at “suture” between Cu2 and 2A (the original description and the original photograph);
4) hindwing slightly angled at M3 (the original photograph; Sato, 2001);
5) abdomen without crests (the original description);
6) labial palpi with moderately long third segment (as in Chlorissa obliterata ) (the original description; Sato, 2001);
7) forewing with R1 (originally “vein 11”) very short, connected with Sc (originally “vein 12”) and dividing the area between Sc and R veins into two halves (the original description); R1 fused with Sc for long distance ( Sato, 2001).
Characters 1-5 are consistent for geometrids in the genera Chlorissa , Nipponogelasma , and Idiochlora Warren, 1896 . Females of all East Asian species of these 3 genera possess labial palpi in which the third segment is very long except for Chlorissa obliterata and Nipponogelasma lucia ( Thierry-Mieg, 1916) , which have a similar, moderately long third segment (character 6). However, Chlorissa obliterata (in contrast to other East Asian Chlorissa species) lacks a green border and white transverse lines, and has a different configuration of the Sc and R1 venation of the forewing, whereas Nipponogelasma lucia has a green border (character 3), and the venation agrees entirely with those described for the holotype of Aoshakuna sachalinensis (character 7). Variation in the Sc and R1 venation in the discussed species is limited: the state described above for the holotypes of Aoshakuna sachalinensis and Nipponogelasma lucia is more common and illustrated by Inoue (1961: fig. 128); the character state with moderately long fusion of Sc and R1 and longer free initial portions of R1 is shown in Fig. 11 View FIGURES 6 – 11 .
Thus, there is little doubt that Aoshakuna sachalinensis is conspecific with Nipponogelasma lucia . As a result, I restore Aoshakuna Matsumura, 1925 , stat. nov., from the synonymy of Chlorissa Stephens, 1831 ; synonymize Nipponogelasma Inoue, 1946 , syn. n., with Aoshakuna Matsumura, 1925 ; synonymize Aoshakuna sachalinensis Matsumura, 1925 , syn. n., with Gelasma lucia Thierry-Mieg, 1916 , with the senior synonym of Gelasma immunis Prout, 1930 , the type species of Nipponogelasma Inoue, 1946 ; and propose the new combination Aoshakuna lucia ( Thierry-Mieg, 1916) , comb. n.
Species composition. The genus Nipponogelasma was monotypic before Inoue (1971) included Mixochlora chlorissoides Prout, 1912 . I consider the latter species to belong to Microloxia Warren, 1893 (see below). Hence, Aoshakuna includes only its type species.
Systematics and phylogeny. The genus Aoshakuna has no closely related genera in East Asia. Inoue (1961) included the genus (as Nipponogelasma ) in the tribe Hemitheini and placed it near Hemithea Duponchel, 1829 . In addition to Nipponogelasma and Hemithea , he included the following East Asian genera in Hemitheini : Chlorissa Stephens, 1831 ; Diplodesma Warren, 1896 (= Idiochlora Warren, 1896 ); Parachlorissa Inoue, 1961 (= Pamphlebia Warren, 1897 ); Culpinia Prout, 1912 ; and Thalera Hübner, 1823 . This generic composition generally corresponds to the tribes Hemitheini and Thalerini sensu Hausmann (2001).
However, Hemithea , the type genus of Hemitheini , has a transtilla that is quite different from that of the other genera – it is a broad plate connected to the valvae by narrow stalks, similar to that found in Jodini, Microloxiini, and Hemistolini (sensu Inoue, 1961, and Hausmann, 2001). Stekolnikov and Kuznetzov (1981) consider this type of transtilla the plesiomorphic state compared to the transtilla of Clorissa, in which this sclerite is solidly connected with the valvae, arched, narrow, and with more developed “lateral lobes.”
This apomorphic type of transtilla is consistent for all genera included in Hemitheini by Inoue (1961) except for Hemithea , and always is accompanied by the presence of a sclerotized dorsal process on the valva. This apomorphy strongly suggests the monophyly of the Palaearctic genera Aoshakuna , Chlorissa , Idiochlora , Pamphlebia , Culpinia , Dyschloropsis Warren, 1895 , and Thalera .
If this hypothesis is correct, Thalerini Herbulot, 1963, should be subordinated to Hemitheini Bruand, 1846 , in the treatment of the latter by Inoue (1961), or in its broader concept, including Jodini Inoue, 1961, following Viidalepp (1981) and Holloway (1996, but as subtribe Hemitheiti). Alternatively, all enumerated Palaearctic genera, except Hemithea , should be transferred to Thalerini.
For North America geometrids, Ferguson (1969, 1985) included in Hemitheini 5 genera: Chlorochlamys Hulst, 1896 ; Xerochlora Ferguson, 1969 ; Chloropteryx Hulst, 1896 ; Hethemia Ferguson, 1969 ; and Mesothea Warren, 1901 . The first three have a wide transtilla similar to that in Hemithea , and the last two possess a transtilla of the Clorissa type. Hence, Ferguson’s treatment of Hemitheini corresponds to its broad concept. For Neotropical Geometrinae , Pitkin (1996) also accepted a broad concept of the tribe.
Epichrysodes Han & Stüning, 2007 , recently was described from the mountains of southeastern China ( Han, Stüning, Xue, 2007), and according to the authors, it exhibits characters of both Thalerini and Hemitheini (sensu Hausmann), supporting the larger concept of Hemitheini . Here I agree with the opinion of previous authors and include Aoshakuna View in CoL in the broad concept of Hemitheini . However, the tribe needs to be evaluated in regards to its possible paraphyly in relation to Hemistolini, Jodini, and Microloxiini.
Within the discussed group of genera, Aoshakuna View in CoL may form a monophyletic group with Epichrysodes , Culpinia View in CoL , Chlorissa View in CoL (for male and female genitalia of East Asian species see Beljaev (1987)), Hethemia View in CoL , and Mesothea View in CoL (for morphology of the last two see Ferguson (1969, 1985)). With them, Epichrysodes , Culpinia View in CoL , and Chlorissa View in CoL (species related to type species of the genus) share a pointed, scobinate dorsal process on the valva, spinules on the apex of the aedeagus, and a W-shaped postvaginal plate, which in Culpinia View in CoL has rudimentary lateral branches (the female of Epichrysodes is unknown). Also, Chlorissa View in CoL and Culpinia View in CoL share a similar apomorphic convex projection on the dorsal side at the middle of aedeagus, which could be an early development of the long scooplike process of the aedeagus present in Aoshakuna View in CoL and Epichrysodes . In Mesothea View in CoL the male has a characteristic knoblike median basal process on the valva that probably is homologous and synapomorphic with that in Chlorissa View in CoL , but the costal process of the valva in the former is smooth, not scobinate. However, a smooth costal process is present in some Chlorissa View in CoL as well (e.g., Chlorissa amphitritaria (Oberthür, 1879)) View in CoL . The structure of the sclerites of the osteal area in females of Mesothea View in CoL is somewhat similar to that in C. amphitritaria View in CoL and suggests a potential close relationship between the two genera. Hethemia View in CoL also may be monophyletic with Chlorissa View in CoL and Mesothea View in CoL based on the female genitalia, which are almost identical with those of Chlorissa View in CoL and possess a putatively synapomorphic setose processes on the postvaginal area. However, the last genus lacks the knoblike median basal process on the valva (probable plesiomorphy) and has the costal sclerite reduced and completely fused with the valva; however, the costal sclerite retains scobination. In addition, the putative close relationship among Epichrysodes , Culpinia View in CoL , Chlorissa View in CoL , Hethemia View in CoL , and Mesothea View in CoL is supported by the presence of a unique scooplike sclerotization on the vesica of the aedeagus; but this sclerotization is absent in Aoshakuna View in CoL (possibly secondarily lost).
In the discussed group of genera, Culpinia may be the earliest derived. Aoshakuna , Epichrysodes , Chlorissa , Hethemia , and Mesothea share the following putative synapomorphies: a conical, spinelike process on the eighth abdominal sternite in the male (bifurcate in Hethemia ); narrow lateral branches (fused caudally in Chlorissa , Hethemia and Mesothea ) derived from the postvaginal plate; and an asymmetric corpus bursae with an inflated posterior wall lateroventrad from the ductus bursae. However, the presence in the male of a conical spinelike process on the eighth abdominal sternite is not unique as it also occurs in some Idiochlora and Microloxia (see below).
The putative sister relationship between Aoshakuna and Epichrysodes is supported by the characteristic mesally-curved socii and the unique, long, posteriorly-directed, scooplike projection on the dorsal side of aedeagus. Unfortunately, the female of Epichrysodes is unknown.
The putative phylogeny of the group is shown in Fig. 21 View FIGURE 21 ; apomorphies are enumerated in Table 1 View TABLE 1 . Relationships among Thalera , Dyschloropsis , Idiochlora , and Pamphlebia are unresolved.
Viidalepp (1981) suspected a sister relationship between Aoshakuna (“ Nipponogelasma ” of the author) and Maxates Moore, 1887 (” Gelasma ” of the author) based on the shared possession of one additional cell between the Sc and R veins of the forewing (character 8 of the author). However, the additional cell occurs in many unrelated genera throughout Geometrinae (clearly shown in fig. 1 of Viidalepp's paper where a phylogenetic tree with plesiomorphic and apomorphic characters are represented), thus the presence of the cell appears to be homoplastic. Other genera from the discussed group included by Viidalepp (1981: fig. 1), i.e., Thalera , Culpinia , and Chlorissa , are distributed on different branches. His phylogeny was based on external features, such as presence/absence of a frenulum, characters of wing venation, and presence of an elongation of the hind tibia in males. The distribution of these characters conflicts with the distribution of the genitalic features. Hence, it is likely that external features are of less value than genitalia for identifying phylogenetic relationships within Geometrinae . Ferguson (1969) emphasized the importance of genitalic characters for Geometrinae systematics and stressed that “loss of the frenulum cannot be assumed to imply relationship” ( Ferguson 1969: 3). I suggest that this statement applies to most other external characters at taxonomic levels above the genus.
Coloration and maculation of adults of Epichrysodes tienmuensis Han & Stüning, 2007 , are quite different from those of Aoshakuna and other members of Hemitheini - the species has a nearly yellow ground color with pinkish-brown maculation. This could be the result of the early spring activity of the imago (March– April) primarily in deciduous high-mountain dwarf forests, when the predominant color of the vegetation is not green. It should be noted that North America species of Nemoria Hübner, 1818 , Chlorochlamys Hulst, 1896 , and Hethemia Ferguson, 1969 , have a brownish spring form ( Ferguson, 1969, 1985) that is unknown in the Palaearctic Geometrinae . Possibly the unusual coloration of Epichrysodes tienmuensis could be the result of a similar adaptation to the cool season.
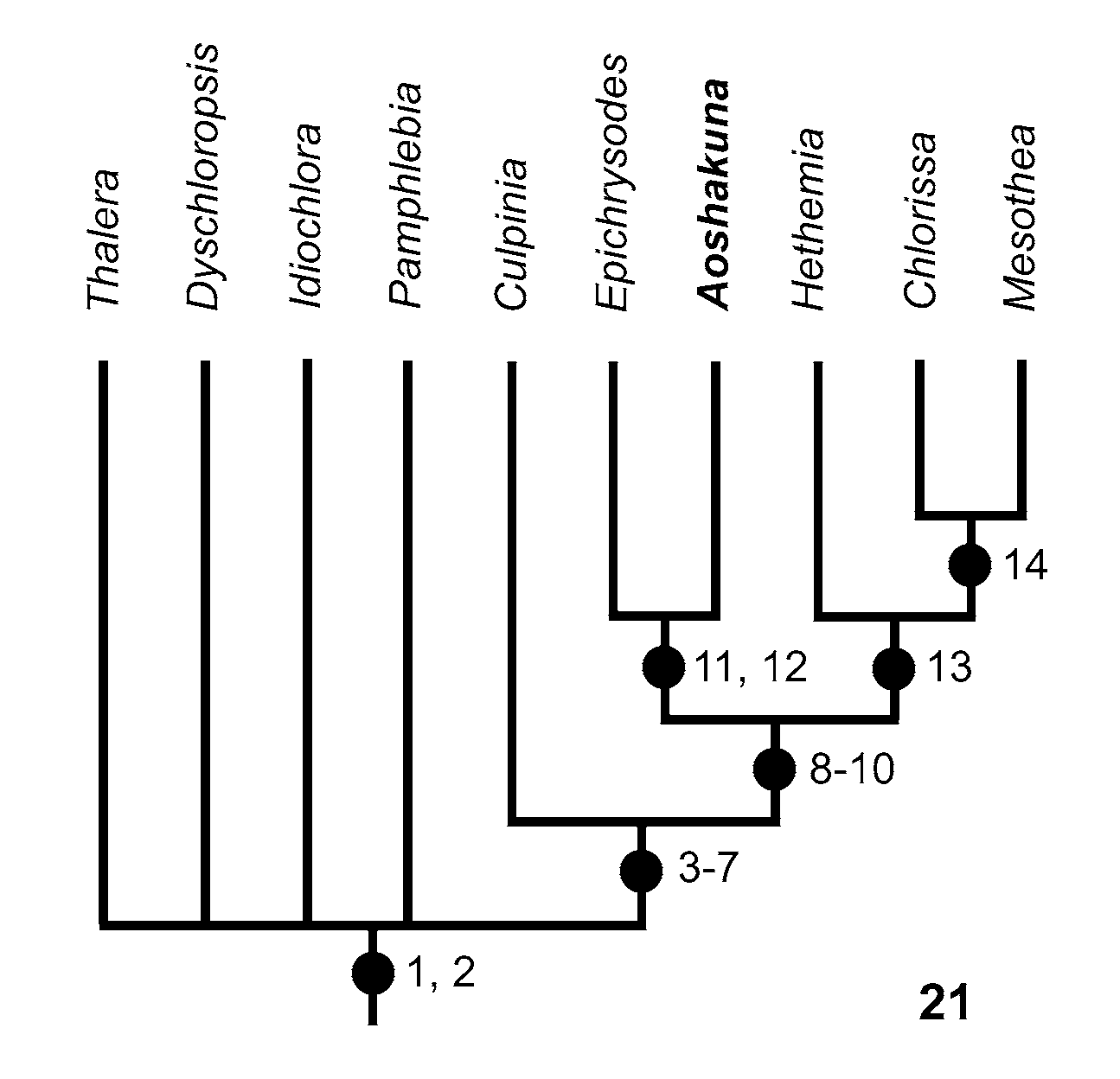


TABLE 1. Apomorphies in the selected genera of Hemitheini.
| Apomorpies | Thalera , Dyschloro- psis, Idio- chlora, Pamphle- bia | Culpinia | Epichryso- des | Aoshakuna | Hethemia | Chlorissa | Mesothea |
|---|---|---|---|---|---|---|---|
| 1. Transtilla solidly con- nected with valvae, narrow, arched, has prominent "lat- eral lobes" | + | + | + | + | + | + | + |
| 2. Dorsal sclerotization on valva with free distal process | + | + | + | + | - (second- arily reduced?) | + | + |
| 3. Dorsal process on valva pointed, scobinate | - | + | + | + | - (scobinate costal bulge) | +/- | - (second- arily modi- fied?) |
| 4. Apex of aedeagus with spinules | - | + | + | + | + | + | + |
| 5. Postvaginal plate W- shaped | - | + | ? | + | + | + | + |
| 6. Aedeagus with convex projection on the dorsal side at the middle | - | + | + (modified, see Ap. 12) | + (modified, see Ap. 12) | ? | + | + |
| 7.Vesica of aedeagus with scoop-like sclerotization | - | + | + (small) | - (second- arily reduced?) | + | + | + |
| 8.Eighth abdominal sternite in male with conic spine-like process | - | - | + | + | + (bifurcate) | + | + |
| 9. Postvaginal plate with narrow lateral branches | - | - | ? | + | + | + | + |
| 10. Corpus bursae asymmet- ric, with inflated posterior wall lateroventrad from duc- tus bursae | - | - | ? | + | + | + | + |
| 11. Socii curved at the mid- dle | - | - | + | + | - | - | - |
| 12. Aedeagus with long, posteriorly directed, scoop- like projection on dorsal side | - | - | + | + | - | - | - |
| 13. Postvaginal aria with pair of setose processes | - | - | ? | - | + | +/- | + (rudimen- tary) |
| 14. Valva with knob-like median basal process | - | - | - | - | - | + | + |
| ZFMK |
Zoologisches Forschungsmuseum Alexander Koenig |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Geometrinae |
|
Genus |
Jodis orientalis Wehrli
| Beljaev, Eugene A. 2007 |
Jodis putata orientalis:
| Parsons 1999: 528 |
Jodis angulata:
| Kim 2001: 30 |
| Shin 1996: 9 |
| Oh 1988: 165 |
| Inoue 1982: 434 |
| Inoue 1977: 234 |