
Kamaka tonlensis, Morino, Hiroshi, 2012
|
publication ID |
https://doi.org/10.5281/zenodo.212565 |
|
DOI |
https://doi.org/10.5281/zenodo.6172863 |
|
persistent identifier |
https://treatment.plazi.org/id/3437166E-FFDB-FFD8-FF4C-FF37CE82FEAA |
|
treatment provided by |
Plazi |
|
scientific name |
Kamaka tonlensis |
| status |
sp. nov. |
Kamaka tonlensis sp. nov.
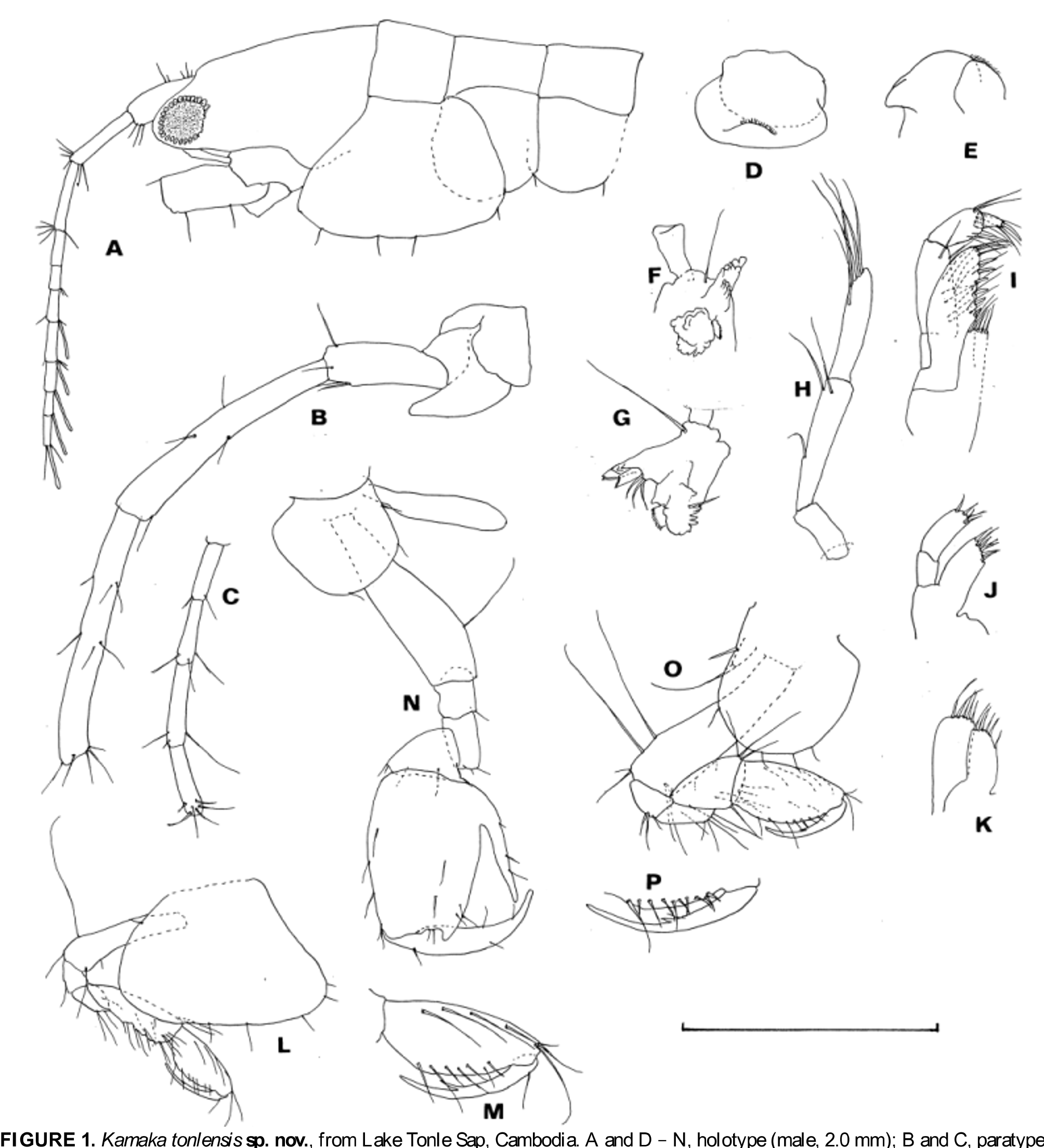
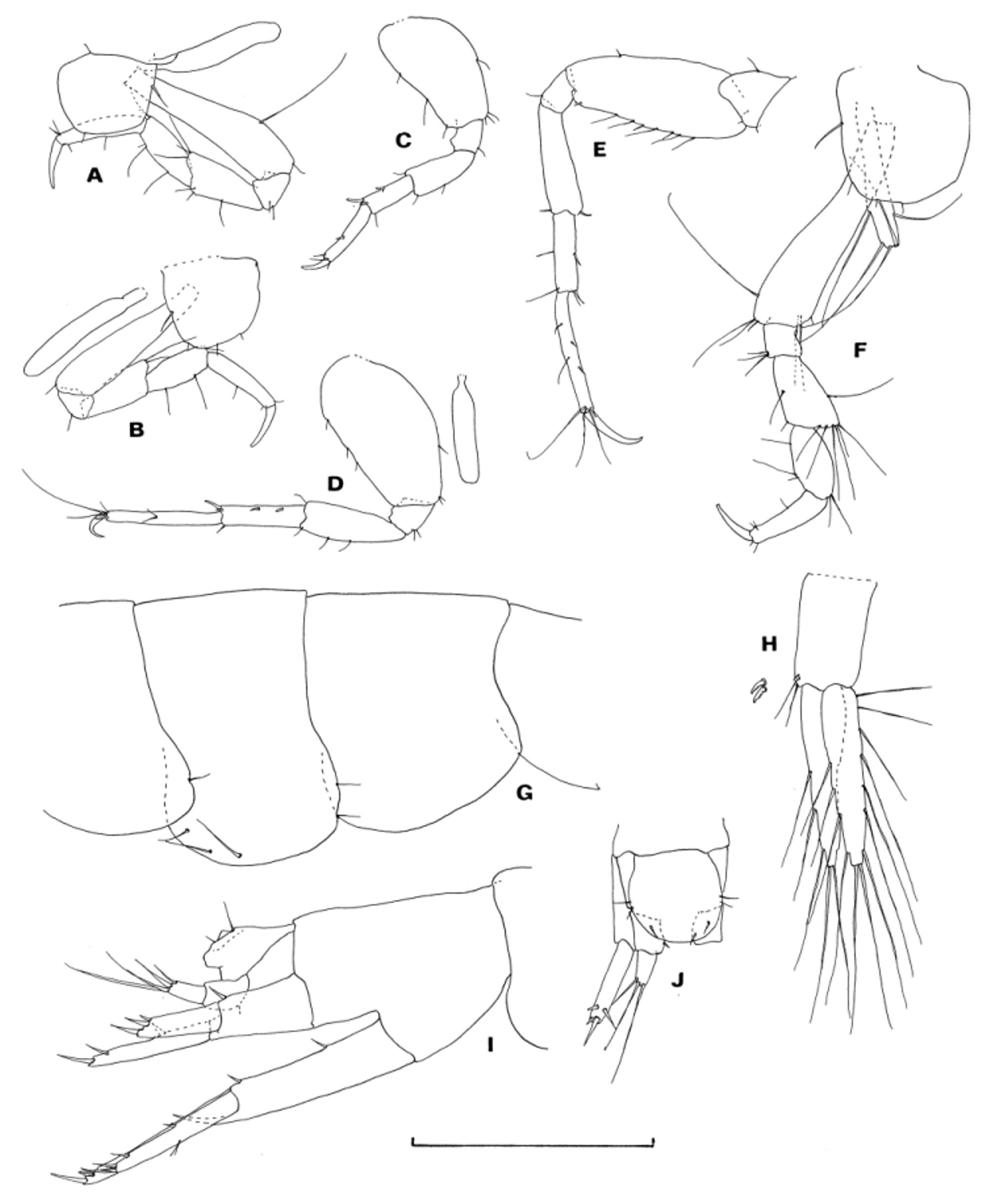
( Figs 1 View FIGURE 1 , 2 View FIGURE 2 )
Material examined. One male ( holotype, 2.0 mm B.L.); South Basin, Lake Tonle Sap, Cambodia, St. 3 ( 12°2’57.7’’N 104°22’04.4’’E, off shore, 4.6 m water depth); 7 December 2004 Ohtaka et al. coll. One female (allotype, 2.1 mm B.L.); South Basin, Lake Tonle Sap, Cambodia, St. 2 ( 12°35’ 58.8”N, 104°20’58.8’’E, off shore, 5.1 m water depth); 7 December 2004, Ohtaka et al. coll. Three males, one female and three juveniles ( paratypes); North Basin, Lake Tonle Sap, Cambodia, St. 1 ( 13°01’54.3”N 103°57’24.3”E, off shore, 0.8 m depth); 13 May 2004, A. Ohtaka et al. coll.
Description. Male. Head ocular lobe ( Fig. 1 View FIGURE 1 A) not reaching tip of peduncular article 1 of antenna 1. Abdominal side plates 1 and 2 ( Fig. 2 View FIGURE 2 G) with a few setae on posteroventral margin, plate 2 with a few setae on ventral margin. Urosomites 1 and 2 ( Fig. 2 View FIGURE 2 I) coalesced. Urosomite 3 ( Fig. 2 View FIGURE 2 I) not coalesced with telson.
Antenna 1 ( Fig. 1 View FIGURE 1 A), ratio of length in peduncular articles 1–3 1.0: 0.9:0.9; flagellum with 7 articles of similar size, distal 5 articles of them with aesthetascs. Antenna 2 ( Fig. 1 View FIGURE 1 B, C), peduncle slender, weakly setose, ratio of length of articles 3–5 1.0: 2.0: 2.1; flagellum with 4 articles. Mandible ( Fig. 1 View FIGURE 1 F–H), basal article with seta near junction with palp; palp articles weakly setose, length ratio of articles 1–3, 1.0: 2.5: 1.9. Other mouth parts ( Fig. 1 View FIGURE 1 D, E, I–K) as other congeneric species.
Gnathopod 1 ( Fig. 1 View FIGURE 1 L, M), coxa posteroventral corner rounded, palmar margin of propodus distinctly longer than the posterior margin, dactylus elongate. Gnathopod 2 ( Fig. 1 View FIGURE 1 N), coxa as deep as wide, propodus posterior margin bluntly produced at distal corner, and with large process proximally, the process not reaching distal tip of posterior margin, dactylus curved inward distally. Pereopods 3 and 4 ( Fig. 2 View FIGURE 2 A, B), basis slender and weakly setose, merus and carpus slender. Pereopods 5–7 ( Fig. 2 View FIGURE 2 C–E), basis weakly expanded, with a few to several setae on posterior margins. Pleopod peduncle ( Fig. 2 View FIGURE 2 H) with a few setae and 2 retinacula on inner distal corner.
Uropod 1 ( Fig. 2 View FIGURE 2 I), peduncle distal process reaching 0.4 length of outer ramus, outer ramus dorsal margin bare, ventral margin with setae at mid-point, and inner ramus with 1 marginal robust seta. Uropod 2 ( Fig. 2 View FIGURE 2 I), outer ramus shorter than inner ramus, outer ramus bare; inner ramus with 1 marginal robust seta. Each ramus of uropods 1 and 2 with 2 subterminal and 2 terminal robust setae, the former 2 are of subequal length, the latter 2 of disproportionately unequal length. Uropod 3 ( Fig. 2 View FIGURE 2 I, J), peduncle with a small spine on posteromedial corner, ramus slightly shorter than peduncle, with 3–4 terminal setae. Telson ( Fig. 2 View FIGURE 2 J) semi-circular, with 2 pairs of setae. Female. Gnathopod 2 ( Fig. 1 View FIGURE 1 O, P), propodus and dactylus similar to those of gnathopod 1 of male. Brood plate rudimentary, with apical seta. Pereopod 3 ( Fig. 2 View FIGURE 2 F), slightly more setose than that of male. Brood plate linear, with 5 setae distally.
Etymology. The specific name refers to the type locality.
Habitat. Kamaka tonlensis has been collected from the bottom of offshore areas of both North and South basins of the lake, irrespective of water level which fluctuates highly with the season ( Ohtaka et al. 2010). The bottom sediment consists of muddy surface layer and the deeper layer of shell remains beneath the surface dominated by Corbicula bivalves (Ohtaka, pers. comm.).
The pH value and electronic conductivity of the Southern Basin (stns 2 and 3) were 7.7 – 7.8 and 86 – 91 μs/ cm at the sampling time, respectively.
Remarks. Kamaka tonlensis sp. nov. is peculiar among congeneric species in having no marginal setae on the outer ramus on the uropod 2 and the relatively elongate dactylus of the male gnathopod 1 (0.83 times as long as propodus). The shape of the male gnathopod 2 of the present new species is exhibited also by K. kuthae Derzhavin, 1923 , K. derzhavini Gurjanova, 1951 , K. littoralis Ren, 2006 and K. silvana Myers, 2009 . Kamaka kuthae and K. derzhavini have the elongate article 1 of flagellum on male antenna 2, by which these species are separable from the new species. Kamaka kuthae is further distinguished from the new species by shorter article 2 of the mandible palp (1.6 times of article 1 vs. 2.5 times), and by more elongate distal process of uropod 1 peduncle (reaching 0.6 length of outer ramus vs. 0.4 length). Kamaka silvana has the longer outer ramus on the uropod 2 than the inner ramus (shorter outer ramus), the urosomite 3 coalesced with the telson (not coalesced), the shorter article 2 on mandiblular palp (1.8 times of article 1) and the setose article 1 (vs. article 1 in the new species lacking setae). Kamaka littoralis has the following features which distinguish it from the new species: the more expanded carpus on the pereopod 3 (width 0.77 times of length vs. 0.47 times), and the enlarged outer plate of the maxilliped that well exceeds the tip of the article 2 of the palp (vs. reaches the tip). In addition, smaller male of K. songkhlaensis Ariyama et al., 2010 displays the similar shape of male gnathopod 2 to that of the present new species (Fig 9 L, Ariyama et al. 2010). The former species is distinguished from congeneric species by expanded peduncular article 4 and excavated flagellar article 1 of antenna 2 in adult male.
Thomas & Barnard (1991) described the urosomite 3 of this genus as “free from (urosomite) 2 but coalesced with telson, marked ventrally by sutures.” This character is not reproduced in the revised diagnosis given by Ariyama et al. (2010). In the recent description of Kamaka species, K. silvana is described to have urosomite 3 coalesced with telson ( Myers 2009). For K. poppi Bamber, 2003 , K. littoralis , K. derzhavini and K. palmata Dang, 1968 , the state of this character is not clear, since no description and no figure are presented for this characters. For the remaining species except K. taditadi Thomas & Barnard, 1991 , no coalescence between the two parts is recognized. In the present new species, there is a clear suture separating the urosomite 3 from the telson ( Fig. 2 View FIGURE 2 I), as that depicted for K. taditadi ( Thomas & Barnard 1991) . However, it is not possible to identify it with an articulation or with a suture covering the coalescence. The description given above is thus a tentative conclusion. In considering the character of fusion of urosomites 1 and 2, which is distinctive and peculiar to this genus, it must be of phyletic importance to recognize the degree or trend of fusion on posterior somites in this group. However, as stated above it seems hard to distinguish the true articulation from the suture on exoskeleton. The revised diagnosis given by Ariyama et al. (2010) may also suggest this impracticability.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |