
Kapateira coffea, Godoy
|
publication ID |
https://doi.org/ 10.5281/zenodo.173429 |
|
DOI |
https://doi.org/10.5281/zenodo.6261015 |
|
persistent identifier |
https://treatment.plazi.org/id/03A6879C-FFFA-FFE4-FED7-EEA4267CFBA9 |
|
treatment provided by |
Plazi |
|
scientific name |
Kapateira coffea, Godoy |
| status |
sp. nov. |
Kapateira coffea, Godoy View in CoL , sp. nov. ( Figs. 1–13 View FIGURES 1 – 7 View FIGURES 8 – 11 View FIGURES 12 a – e View FIGURES 13 )
Type material: Holotype: male, COSTA RICA: San José: Desamparados, 1200m, 5I 2001, C.M. Rodríguez ( CAS). Paratypes: 13 males and 6 females, same data as holotype; 4 male and 7 female, same data except 19I2000; 1 female, same data except 8II2000; 2 males and 3 females, same data except 15II2000; 4 males and 7 females, same data except 21XII2000; 2 females same data except 8II2001; 4 males and 3 females, same data except 12II2001; 2 females, same data except 8III2001; 1 male and 2 females, same data except 19III2001; 1 male and 3 females, same data except 28III2001; 1 male and 2 females, same data except 6IV2001; 2 females, same data except 21XII2001; 1 male and 3 females, same data except 29X2001 ( CAS, INBio, BMNH, UCR).
Description. Length of male 6.7–7.2 mm, female 6.5–7.4 mm. General color mustard yellow, with black spots or lines on the dorsum of head; pronotum with two spots ( Fig. 1 View FIGURES 1 – 7 ).
Male genitalia: pygofer with ventral posterior margin rounded, posteriorly produced into short spine on dorsal posterior margin ( Fig. 2 View FIGURES 1 – 7 ). Subgenital plate narrowly triangular, extending posteriorly beyond apex of pygofer, with uniseriate macrosetae ( Figs. 2, 3 View FIGURES 1 – 7 ).
Style extending posteriorly beyond apex of connective, without preapical lobe ( Fig. 5 View FIGURES 1 – 7 ). Connective short, Yshaped ( Fig. 4 View FIGURES 1 – 7 ). Aedeagus symmetrical in lateral view with a paired elongate apical processes projecting distally, gonopore located apically, atrium with spine, basal apodemes greatly elongate ( Figs. 6, 7 View FIGURES 1 – 7 ).
Female sternum VII with ventral surface convex, posterior margin pointed at apex and concave on both sides ( Fig. 8 View FIGURES 8 – 11 ). First and second valvulae elongate, slightly wider subapically, tapered to apex. First valvula in lateral aspect wider in apical half and slightly curved, with strigate sculpture on dorsal and apical margins ( Fig. 9 View FIGURES 8 – 11 ). Second valvula in lateral aspect with dorsal teeth, individual teeth rounded and slightly elevated ( Fig. 10 View FIGURES 8 – 11 a–b). Third valvula shorter, narrowed apically to rounded point ( Fig. 11 View FIGURES 8 – 11 ).
Nymphs ( Figs. 12 a–e View FIGURES 12 a – e ). Color yellowish, with four distinct longitudinal brown to black stripes. Head large, as long as basal width, crown with anterior portion brown to black; clypeus broad with two longitudinal stripes. Abdominal tergites VI to VIII each with short setae on posterior lateral margins, usually more setae on tergite IX. There are five instars, each resembling the other except for size and the developing wings as shown in Fig. 12e View FIGURES 12 a – e . The developing wings, in the form of wing pads, become more pronounced in the third, fourth and fifth instars. In these same instars the tibiae have a brown line between setal rows I and II. The nymphs vary from 1.6 to 1.8 mm in length in the first instar, to 6 mm in the fifth instar.
Distribution. Known only from Costa Rica.
Diagnosis. Kapateira coffea is similar to K. rosipennis but the pygofer has the ventral margin rounded and it is produced into a short spine. Moreover, the aedeagus has paired processes, which is very distinct from the aedeagus of K. rosipennis .
Etymology. The specific name refers to the host plant Coffea arabica.
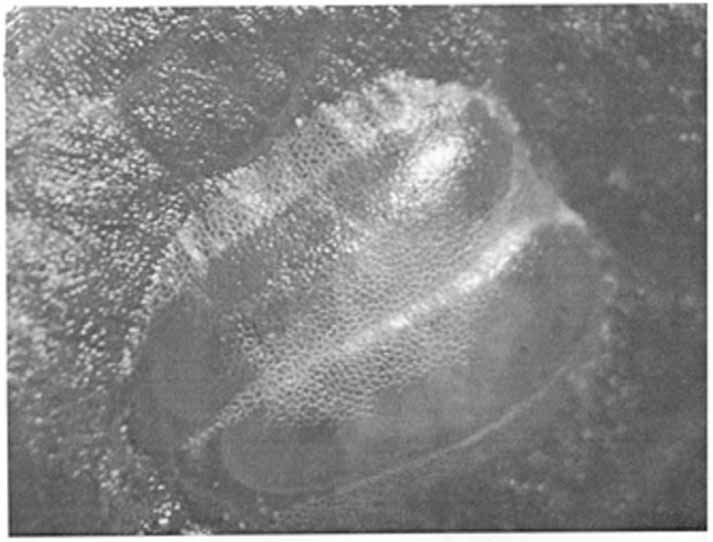
Biology. The eggs ( Fig. 13 View FIGURES 13 ) are generally laid in pairs, rarely individually, the two eggs together and in the same position. They are covered with a transparent mucilaginous secretion and are inserted just under the surface of the epidermis, on both the underside and the upperside of the leaf, occasionally on young, nonwoody stems. Each egg is about 2 mm long and yellow. They are fairly difficult to find even with a hand lens, but can be recognized as small, beanshaped blisters ( Fig. 13 View FIGURES 13 ). The red eye of the developing nymph can be recognized shortly before it hatches. The egg stage lasts about 7 days (n = 10).
Immediately after eclosion the nymphs began feeding, generally on the underside of young leaves. The average duration of the first stadium was 11 days (n=10). Nymphs observed in the process of molting in the laboratory (n=10), were always located on the undersides of mature leaves on the lower part of the plant. The average duration of the second, third, and fourth stadia was 5 (n=9), 4.5 (n=8), and 9.5 days (n=8), during which time feeding behavior remained the same, except that the fourth and fifth instars began feeding for greater lengths of time on nonwoody stems. The average duration of the fifth stadium was 11 days (n=6). Of the ten total individuals under observation in the laboratory, one died during the second stadium, one in the third, and two died during the fourth stadium.
The adult stage, which lasted about 37.5 days (n=6), fed on both young and mature leaves. Adults also fed on young stems, on which they remained for most of the day. A total of 120 individuals were tested with DASELISA, and of these 43 were positive for Xylella fastidiosa . This suggests that K. coffea is a potential vector of plant diseases caused by this bacterium, although further tests are needed to determine its actual role in disease transmission. In North America X. fastidiosa has a diverse host range encompassing over 30 families of monocotyledonous and dicotyledonous plants ( Purcell and Hopkins, 1996). In Costa Rica this bacterium causes coffee leaf scorch and citrus variegated chlorosis, although the latter seems to occur primarily when citrus is planted in coffee plantations ( Rodríguez, et al. 2001; Aguilar et al. 2005).



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Cicadellinae |
|
Genus |