
LITTHABITELLIDAE Falniowski and Hofman, 2023
|
publication ID |
https://doi.org/10.1080/00222933.2023.2168573 |
|
DOI |
https://doi.org/10.5281/zenodo.7778208 |
|
persistent identifier |
https://treatment.plazi.org/id/C92D878F-3C66-D354-255D-FB3DFE30FD64 |
|
treatment provided by |
Plazi |
|
scientific name |
LITTHABITELLIDAE Falniowski and Hofman |
| status |
fam. nov. |
Family LITTHABITELLIDAE Falniowski and Hofman View in CoL , fam. nov.
urn:lsid:zoobank.org:act:A8E1F6A7-EEAE-4B8D-A82D-19A7144283C6
Type genus
Litthabitella Boeters, 1970 View in CoL
Diagnosis
Minute truncatelloid snails with the following synapomorphic character states: osphradium broadly ovate; pallial oviduct gland complex short; gonoporus deep inside the mantle cavity; the ventral channel not on the ventral side of the capsule gland, but somewhat lateral; thickened oviduct circular; the distal termination of the penis wide and blunt, with sharp and short filament harbouring the vas deferens situated laterally, and with two flat, cockscomb-like, four-folded lobes.
Description
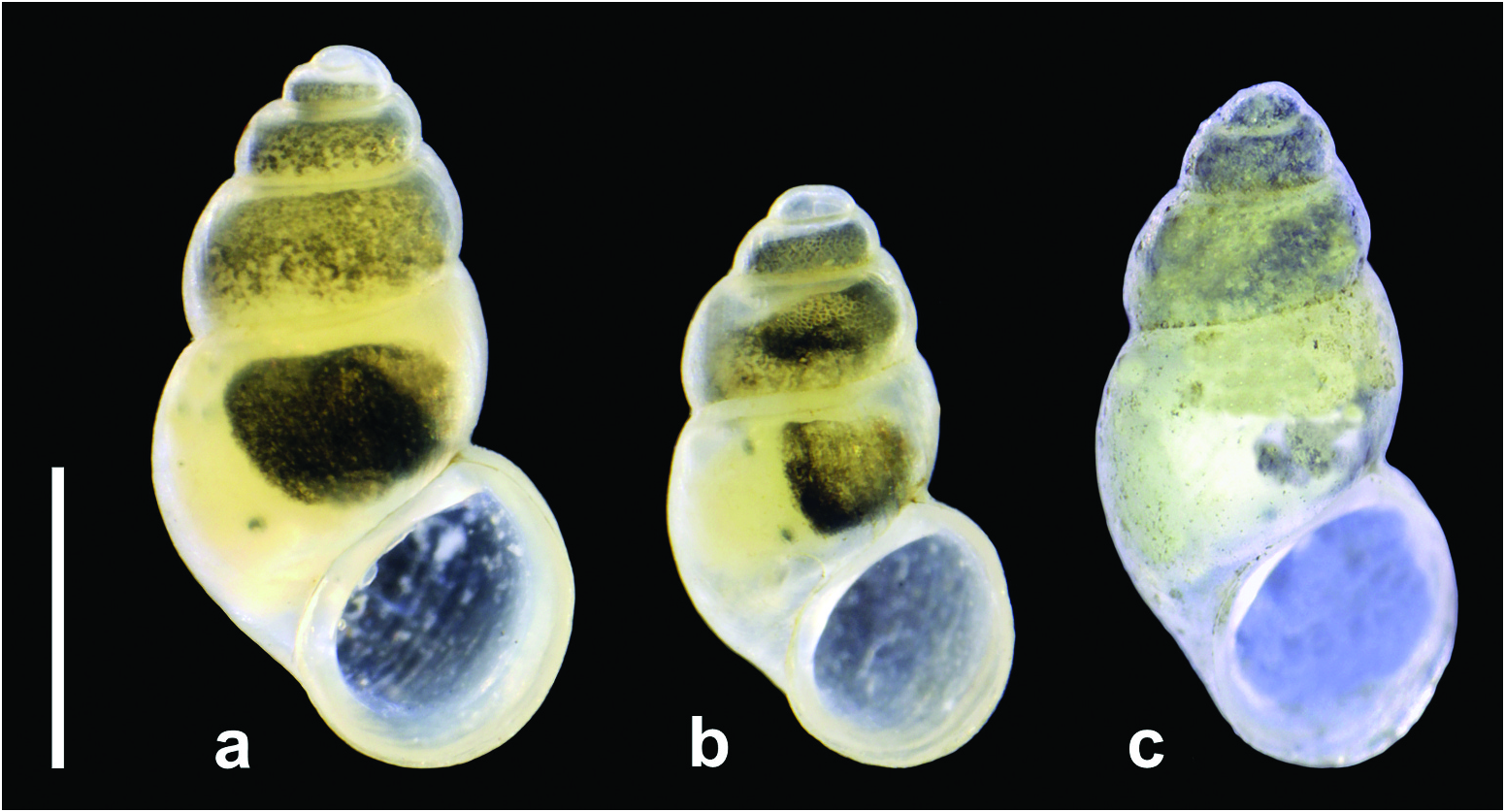
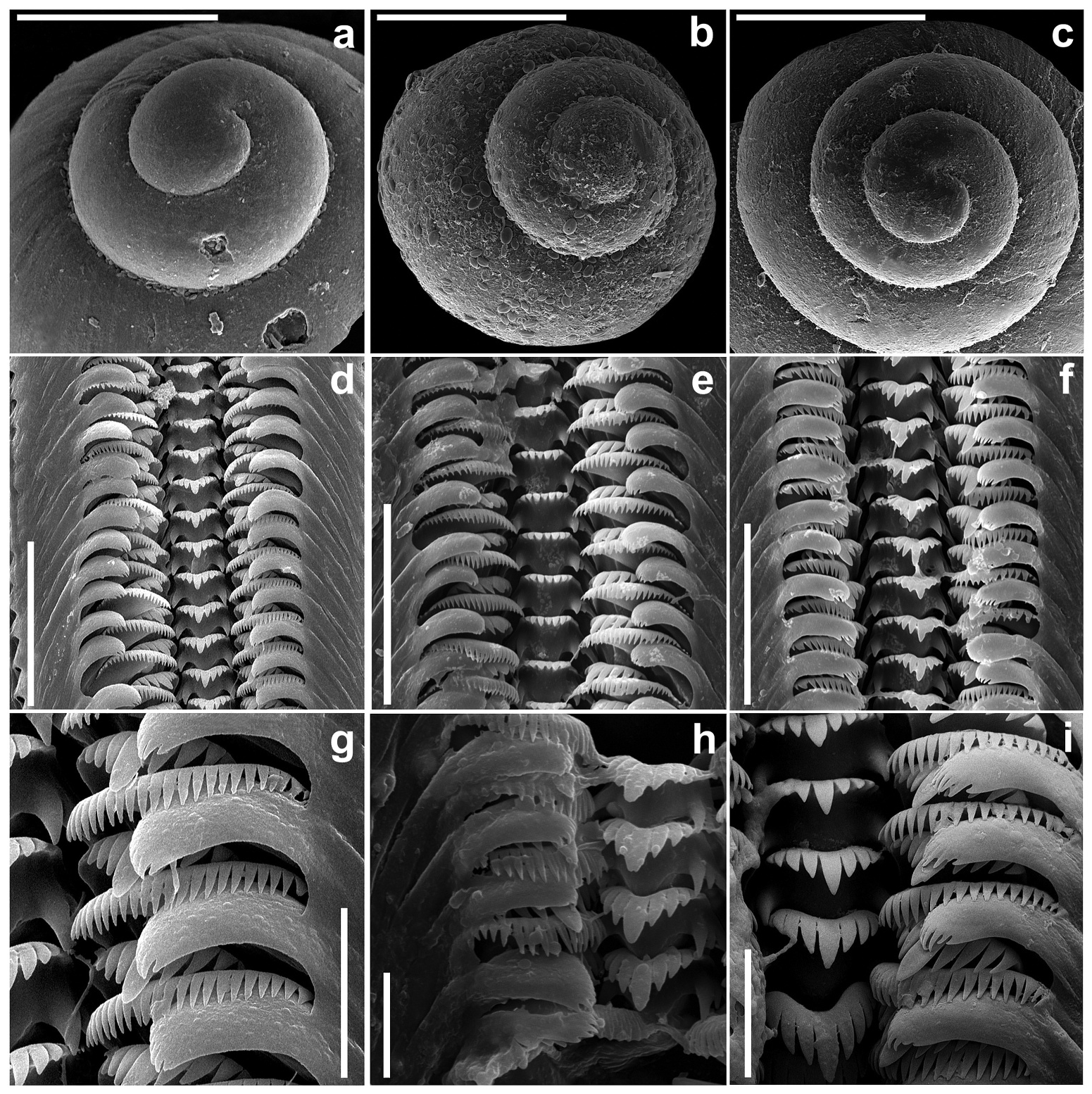
Shell ( Figures 3 View Figure 3 and 4 View Figure 4 ) ovate conical, convex in outline, thick-walled; the border between the proto- and teleoconch is easily discernible, the initial part of the protoconch narrow, on the protoconch surface only delicate irregularities ( Figure 5 View Figure 5 (a–c)), aperture pyriform, teleoconch smooth with delicate growth lines. Operculum ( Figure 6 View Figure 6 ) thin and horny, colourless and translucent, elongate-ellipsoidal, spiral, paucispiral, with submarginal nucleus. Head and tentacles white, with no pigment, no hyperciliation (long and dense cilia at one or two cephalic tentacles, characteristic of e.g. some Hydrobiidae ); mantle partly black pigmented. Osphradium ( Figure 7 View Figure 7 (a)) broadly ovate; ctenidium well developed.
In the radula ( Figures 5 View Figure 5 (d–i) and 8(b)) the central tooth with deep sinus at the proximal part of the cutting plate and one pair of large and prominent basal cusps. The median cusp narrow and slender, more than twice as long as the adjacent cusps, similar in shape. On each side of the median cusp are four to five cusps; the cutting edge is straight, with no cusps. In the lateral tooth, the biggest cusp is triangular, twice as long and broad as the adjacent ones. The formula of the tooth is (5)4–1–5; on the inner marginal tooth 26–28 long and slender cusps. The caecum is absent ( Figure 8 View Figure 8 (c)).
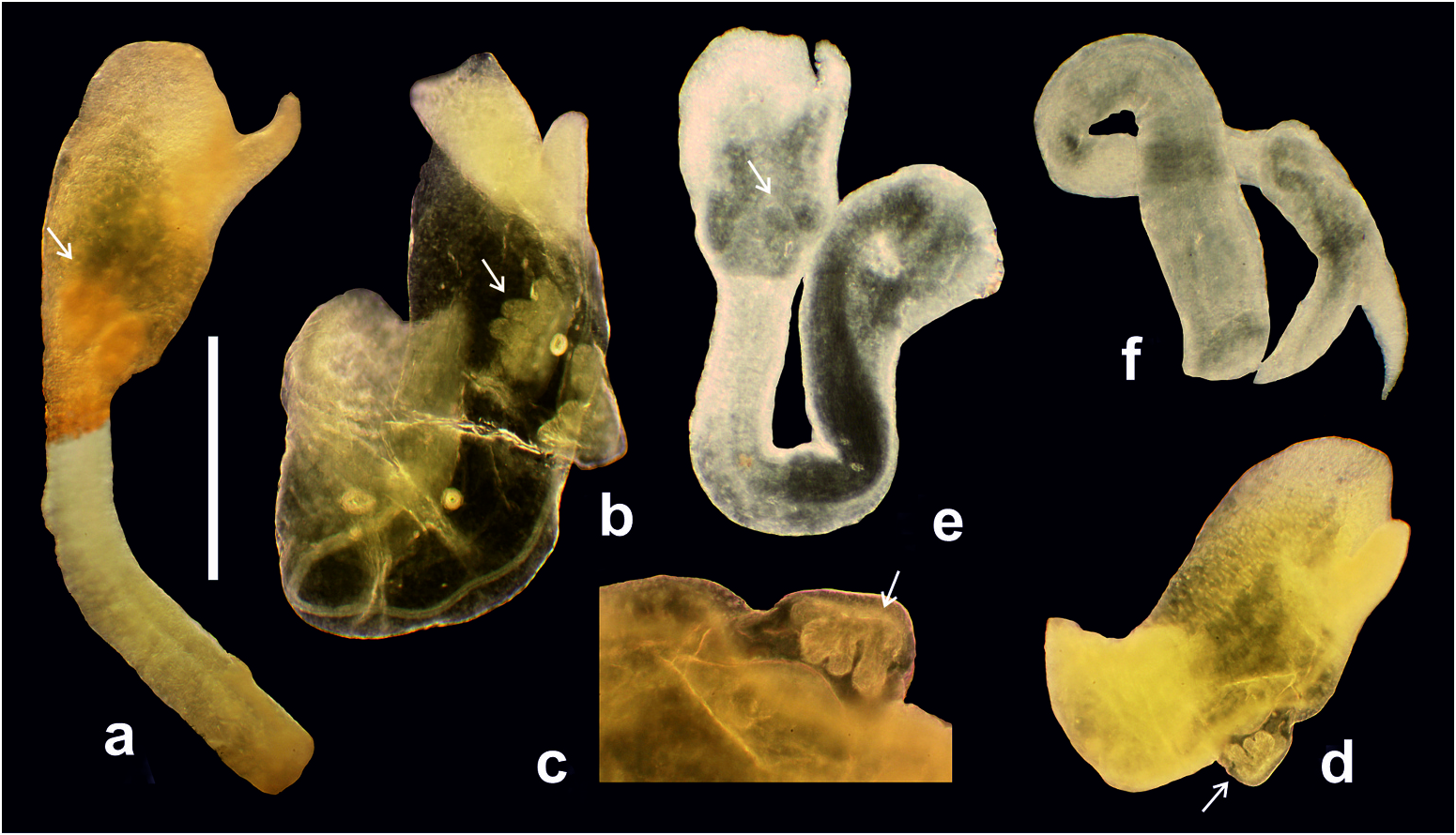
The pallial oviduct gland complex ( Figure 7 View Figure 7 (a)) is relatively short, lying far posteriorly of the mantle margin, thus the gonopore is at the terminal part of the capsule gland and situated deep inside the mantle cavity, far behind the anus. The ventral channel is not on the ventral side of the capsule gland, but lies somewhat laterally ( Figure 7 View Figure 7 (b,d)). There is a bursa copulatrix and two receptacula typical of the Truncatelloidea : a distal one (rs 1 after Radoman 1983) situated near the junction of the oviduct and the duct of the bursa, and a proximal one (rs 2 after Radoman 1983) situated at the proximal end of thickened oviduct; in Litthabitella the proximal receptaculum is vestigial. The thickened oviduct is circular, forming neither a spiral nor a horseshoe-like loop ( Figure 7 View Figure 7 (c)). The prostate is small, nearly vestigial, observable as slight outgrowth of the vas deferens. The penis is big and massive ( Figures 9 View Figure 9 and 10 View Figure 10 ), filled with glandular tissue ( Figure 10 View Figure 10 ), with a distal termination that is wide and blunt, with a sharp and short filament harbouring the vas deferens, which is situated laterally, and two flat, cockscomb-like, four-folded lobes.
Molecular phylogeny
We obtained 26 new sequences of COI (457 bp, GenBank accession numbers ON661272–ON661297), 16 of H3 (310 bp, GenBank accession numbers ON661565– ON661580) and 24 of 18S (401 bp, GenBank accession numbers ON667998– ON668023). The tests by Xia et al. (2003) revealed no saturation. Results from the substitution saturation analysis showed an Index of substitution saturation (ISS) (0.71 for COI; 0.48 for H3) significantly smaller than the critical ISS value (ISSC: 0.97 for COI; 0.59 for H3), indicating that all sequences are useful in phylogenetic reconstruction. In all analyses, the topologies of the resulting phylograms were identical in the ML and BI phylogram analyses; thus, we present the phylogram computed with RAxML.
The COI phylogram ( Figure 11 View Figure 11 ) clearly shows all the populations of Litthabitella as a distinct, highly supported group (bootstrap 99%). As usual with COI, deeper nodes are not well supported; thus, the relationships among the Litthabitella remain unresolved. In the 18S phylogram of Szarowska (2006), Emmericia was a sister clade to all the Hydrobiidae (including Hydrobiinae and Sadlerianinae sensu Szarowska 2006). The time of divergence of the Litthabitellidae was estimated as 8.69 ± 1.87 Mya in the COI phylogram.
Our 18S phylogram ( Figure 12 View Figure 12 ) shows a similar picture: a well-supported clade (bootstrap 83%) includes all the Hydrobiidae and Litthabitella , with the latter as the sister clade of all the Hydrobiidae . The set of available sequences of histone H3 was very restricted, but the relationships ( Figure 13 View Figure 13 ) are similar to those in the 18S phylogram; the well-supported clade (bootstrap 97%) includes Litthabitella as a sister clade of all the Hydrobiidae , but also of Heleobia (Cochliopidae) . The phylogram inferred for the three concatenated loci ( Figure 14 View Figure 14 ) also shows the Litthabitellidae as a sister taxon to all the Hydrobiidae , but the clade joining the Litthabitellidae and Hydrobiidae is not well supported (only 62%); the clade Litthabitellidae is well supported (bootstrap 100%).









No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Caenogastropoda |
|
Order |
|
|
SuperFamily |
Truncatelloidea |
|
Family |