
Lophuroecia lophura ( Müller, 1906 ) Chavtur & Bashmanov, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4516.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:47238257-4DC1-4CF3-A07F-862FFD5E4ECF |
|
DOI |
https://doi.org/10.5281/zenodo.5959295 |
|
persistent identifier |
https://treatment.plazi.org/id/03DB87F4-1F1E-2C6D-10ED-FC3AF538FBA5 |
|
treatment provided by |
Plazi |
|
scientific name |
Lophuroecia lophura ( Müller, 1906 ) |
| status |
comb. nov. |
Lophuroecia lophura ( Müller, 1906) View in CoL comb. nov.
( Figs. 29–32 View FIGURE 29 View FIGURE 30 View FIGURE 31 View FIGURE 32 )
1906 Conchoecia lophura —Müller: 99, pl. XX, figs. 1-10;
1920 Conchoecia lophura —Skogsberg: 689, fig. CXXXI;
1968 Conchoecia lophura —Deevey: 89, fig. 44;
1973 Conchoecia lophura View in CoL — Poulsen, 1973: 125 –128, fig. 61;
1979 Conchoecia lophura View in CoL lissoides— Martens, 1979: 323 –327, Abb. 10 a–d, 11 a–g, Taf. II E, F;
1980 Conchoecia lophura View in CoL — Deevey& Brooks, 1980: 107, fig. 33; Angel, 1993: 176, fig. 62;
2012 Conchoecia lophura View in CoL — Drapun & Smith, 2012: 54 –57, pict. 6, pls. 15, 16, figs. 18, 19.
Examined material. FT Pelamida, 25 th Cruise: MIMB 18349/1—adult male (2.63 mm), 18349/4—adult female (2.94 mm), station 58, sample 23, 38°00´N – 149°00´E, layer 800– 600 m, May 29, 1974. RV Vityaz, 57 th Cruise: MIMB 18349 View Materials /2—adult male (2.61 mm) and 18349/5—adult female (2.78 mm), station 7236, sample 34, 5°57´1´´– 6°00´0´´ N and 123°40´1´´– 124°01´1´´ E, layer 750– 500 m GoogleMaps , sounding 4760– 3830 m, 26–27 February 1975. RV Akademik Mstislav Keldysh, 22nd Cruise: MIMB 18 View Materials 349/3—adult male (2.70 mm) and18349/6—adult female (2.90 mm), station 2345, sample 98, 29°9.36´N – 141°16.21´W, layer 200– 100 m GoogleMaps , sounding 4900 m, September 12, 1990.
Additional material. See Appendix.
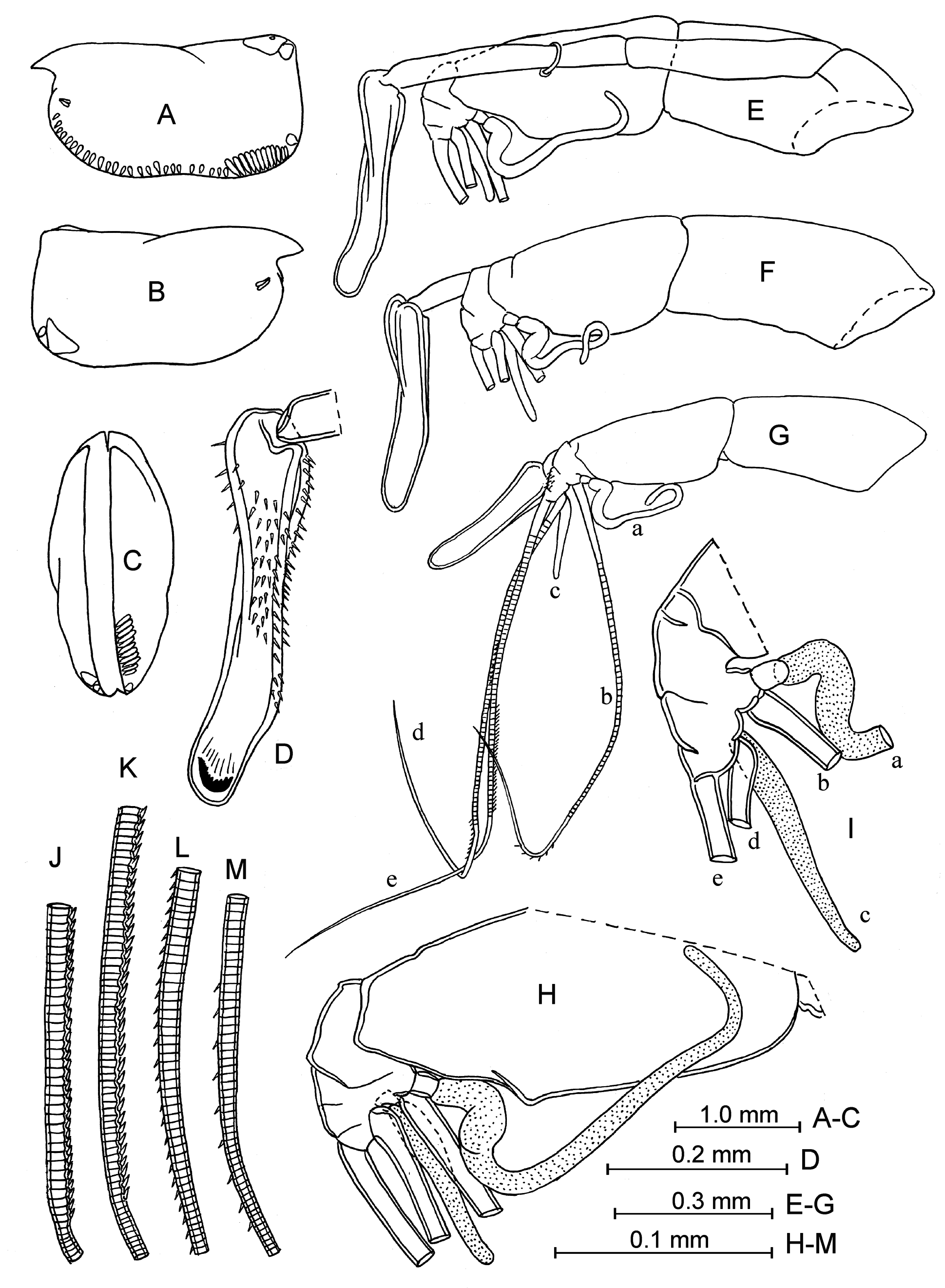
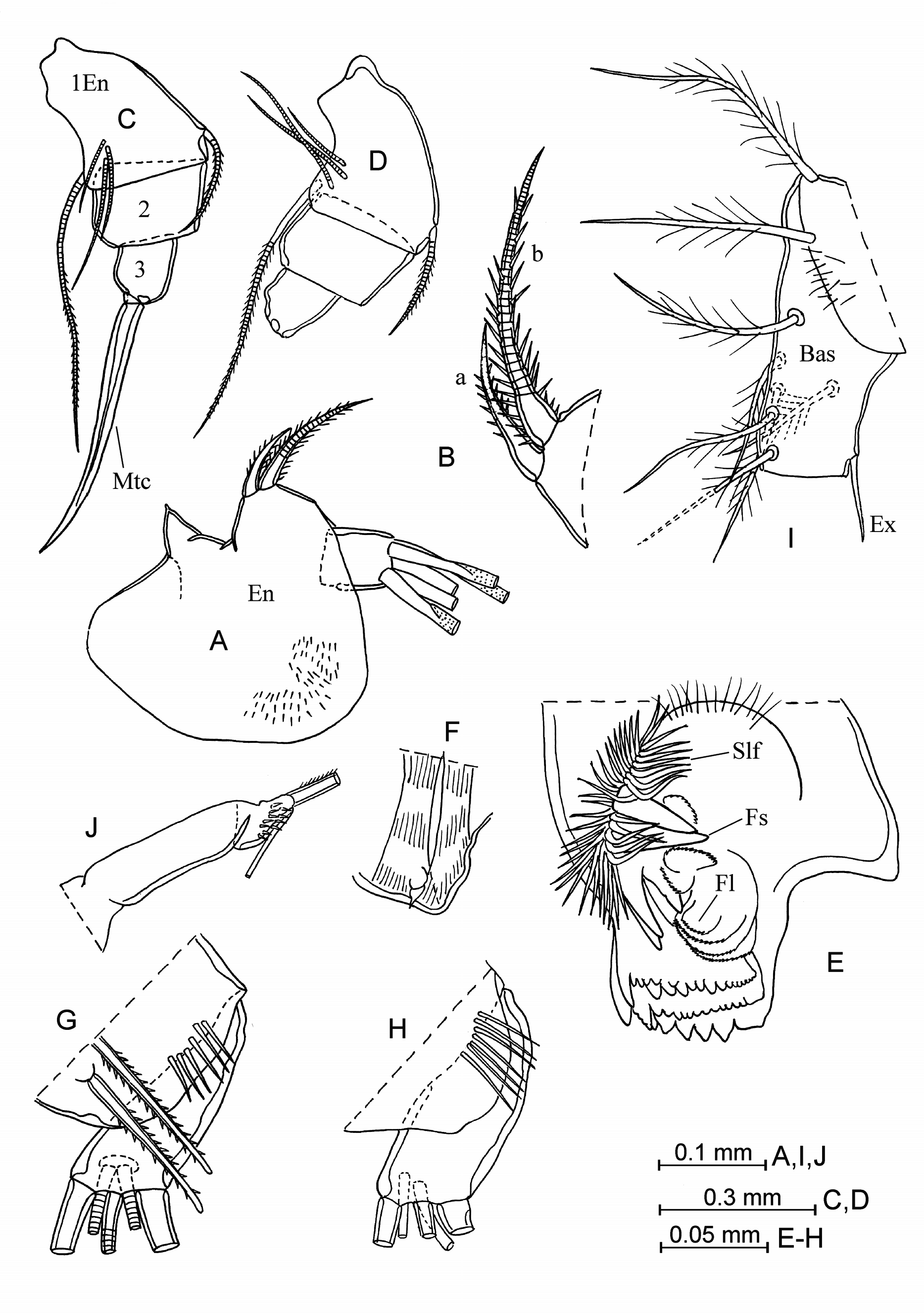
Addition description of adult male. Carapace ( Fig. 29 View FIGURE 29 A–C). Length is between 2.55–2.70 mm at the northwestern Pacific and 2.65–3.00 mm at the north-eastern Pacific (in the literature from 2.08 mm in Drapun & Smith 2012 to 2.84 mm in Martens 1979). The lateral outline is sub-rectangular, slightly tapered (summary literature data from Müller 1906; Skogsberg 1920; Angel 1993; Blachowiak-Samolyk & Angel 2004: may or may not be tapered). Greatest height is 50–52% at the posterior parts of the carapace. The posterior margin is almost straight or slightly arched, and the ventral margin at most with a slight concavity. The right asymmetrical gland opens at the postero-ventral corner, and the left asymmetrical gland is positioned slightly anteriorly. Each valve has a single lateral gland near the postero-ventral corner, and another one near the postero-dorsal corner. There are gland cells beneath the rostral incisures. The left valve has a series of closely spaced long and narrow gland cells at the postero-ventral corner. Surface of the carapace is sculpted with concentric lines, which are distinct in our specimens but are reported to be indistinct in other material Drapun & Smith (2004).
Frontal organ ( Fig. 29 View FIGURE 29 D–G). It is bent downwards. The capitulum is relatively slim in the north-western Pacific but broader in the north-eastern Pacific, but consistently with a slight dorsal concavity. The capitulum end is rounded and slightly down-turned. Surface of the capitulum is well covered with spines.
First antenna ( Fig. 29 View FIGURE 29 E–M). Lengths of the first and the second segments are similar. Seta-a extends just beyond the suture between the first and the second segments of the limb. Seta-b has few (usually about six or nine) small spines and seta-d is armed with nine or more similar spines opposite the comb of seta-e (specimens from the north-eastern Pacific have fewer spines than those from the north–western Pacific). Seta-c is slightly longer than the combined lengths of the third, fourth and fifth segments. Armature of seta-e consists of a comb with about 70– 75 pairs of small and directed proximally spines (summary literature data from Skogsberg 1920; Martens 1979; Angel 1993; Blachowiak-Samolyk & Angel 2004; Drapun & Smith 2012: 60–71 pairs; M.V. Angel’s private message: from 38–42 pairs in specimens from the Atlantic and Banda Sea and up to 80 in specimens from the north Pacific); the length of spines are shorter than the diameter of the seta. Surfaces of the first and the second segments are bare.
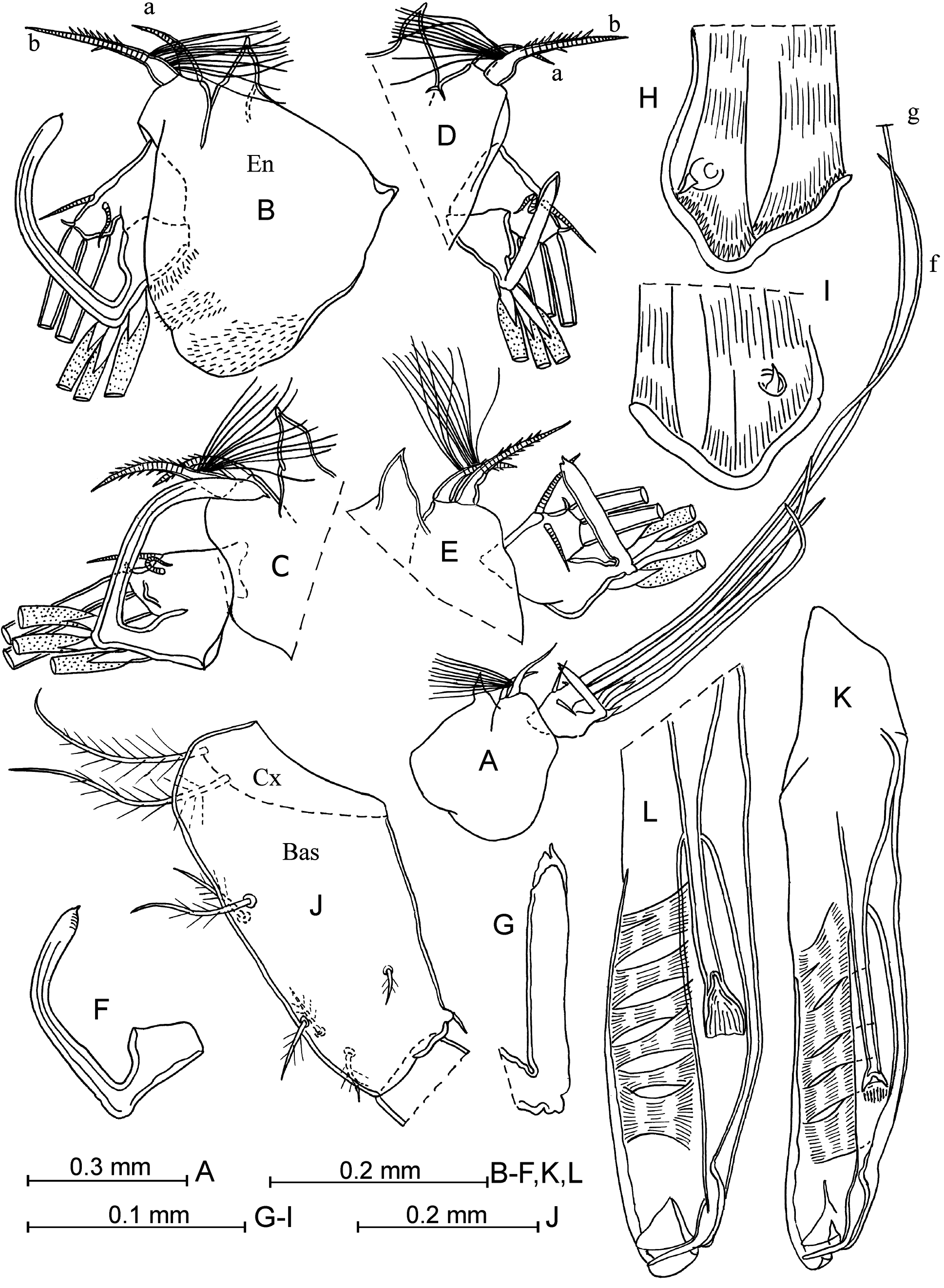
Second antenna ( Fig. 30 View FIGURE 30 A–G). Seta-b of the endopodite has about eight long, basal, fine filaments in the northwestern Pacific and 12–14 filaments in the north-eastern Pacific (summary literature data from Müller 1906; Skogsberg 1920; Martens 1979; Drapun & Smith 2012: about 6–20). The right clasping organ is sub-squared or somewhat oblique proximally, unswollen, and with a rounded end armed with a terminal spine. The left clasping organ is moderately thick, untapered and right- or acutely-angled, with a pointed or spine-like tip. Setae-h–j vary in length, and are relatively slim, slightly tapered towards to the end. Seta-g is about 2–2.5 times as long as the setae-h–j.
Mandible ( Fig. 30 View FIGURE 30 H–I). The epipodite has a rudimentary seta on the verruca. The ventral margin of the first endopodite segment bears four setae (one long seta and three medium-length or short setae). The disto-dorsal seta of this segment is plumose. The toothed edge of the coxale endite is armed with about eight teeth, and the distal tooth-list and proximal tooth-list have 15 and 19–20 teeth respectively (in Poulsen 1973: 6, 12 and 15 teeth). The masticatory pad has four rounded flaps, four flat, long spines and one small, sclerotized tubercle (or short spine?) and about 31–42 seta-like filaments.
Maxilla. The first endopodite segment has six anterior setae and three posterior ones. Anterior setae are in typical locations. Along the distal edge of this segment is a line of five or six long spines.
Fifth limb. The basal segment has a proximal group of three or four setae ventrally, a medio-lateral group of two and a distal group of three setae. The basal segment has a lateral and dorsally long seta (vestige of the exopodite), which extends beyond the end of the limb. The first endopodite segment bears two ventral setae and one dorsal seta.
Sixth limb ( Fig. 30J View FIGURE 30 ). The coxale has two long, plumose setae. The basale bears five ventral plumose setae (one long and one short, proximal setae, and three short, distal setae) and one lateral short, plumose seta. The exopodite slightly extends beyond the proximal boundary of the first endopodite segment.
Seventh limb ( Fig. 32J View FIGURE 32 ). The distal segment has about 11–13 long thin spines.
Caudal furca. An unpaired seta is present.
Copulatory appendage ( Fig. 30K, F View FIGURE 30 ). It is elongated, broader at the middle part, and tapered to a rounded end. The distal seta is thin. The appendage is large, triangular and with a rounded tip. The limb has five or six oblique muscle bands.
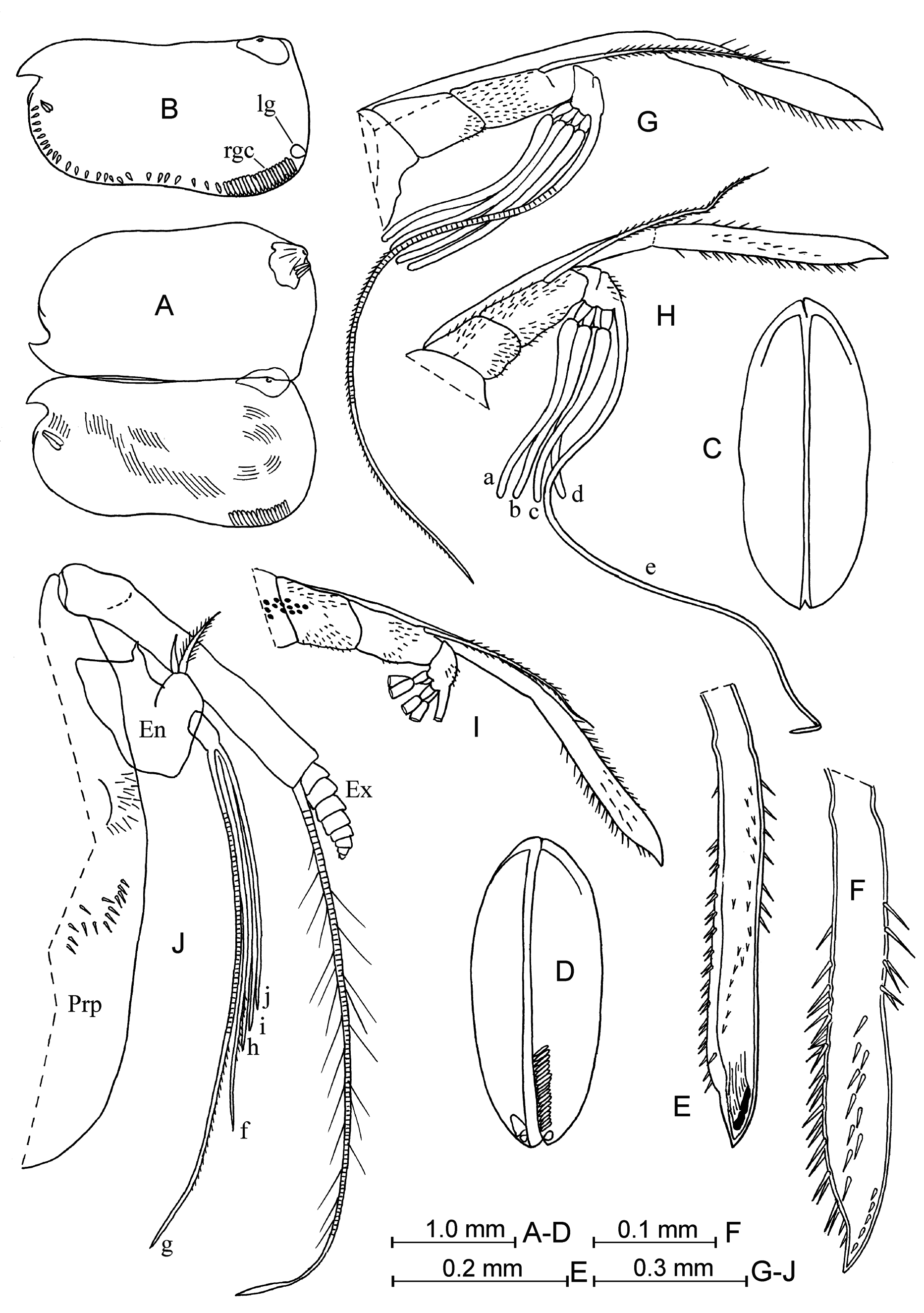
Addition description of adult female. Carapace ( Fig. 31 View FIGURE 31 A–D). Length is between 2.75–2.95 mm at the north-western Pacific and 2.45–2.75 mm at the north-eastern Pacific (in the literature: from 2.20 mm in Müller 1906 to 3.08 mm in Martens 1979). The height is 53–57%. It tapers anteriorly and has lower shoulder vaults than those in the male. The posterior margin is arched, and the ventral margin is concave. Asymmetrical and lateral corner glands have locations usual for the genus. Sculpture is similar to that of the male.
Frontal organ ( Fig. 31 View FIGURE 31 E–I). The stem is slightly bent down. The capitulum is fused with the stem, straight, somewhat thicker at the distal half, with a pointed and slightly down-pointed end. The capitulum surface is almost completely covered with long, thin spines.
First antenna ( Fig. 31 View FIGURE 31 G–I). The dorsal seta on the second segment is well developed, finely spinouse and reaches to about 1/3–3/4 the length of the capitulum ( Angel 1993: in the Atlantic it barely reaches level with the proximal end of the capitulum).Setae-a–d are as long as than the shaft of the limb. Seta-e is proximally bare on its anterior margin, slightly tapered towards a pointed end. The surface of the limb is covered with small spines.
Second antenna ( Figs. 31J View FIGURE 31 ; 32A, B View FIGURE 32 ). The first endopodite segment is about 29%, 40% and 53% the lengths of setae-g, -f and –h, respectively in our specimens (25%, 34% and 42% in Drapun and Smith 2012). All terminal setae on the endopodite are of similar thickness through their lengths and have pointed tips. Seta-b on the first endopodite segment has a swollen base and is armed with obvious stiff spines.
Mandible ( Fig. 32 View FIGURE 32 C–F) and caudal furca are similar to those of the male.
Maxilla (fig. 32G, H). The limb is the same as in the male except there are six or seven spines along the distal edge of the first endopodite segment.
Fifth limb. As in the male.
Sixth limb ( Fig. 32I View FIGURE 32 ). The coxale has two long, plumose setae. The basale bears five ventral setae and one lateral seta; all are long and plumose. The exopodite extends well beyond the proximal suture of the first endopodite segment.
Remarks. Our specimens differ from the descriptions in the literature ( Skogsberg 1920; Martens 1979; Angel 1993; Blachowiak-Samolyk & Angel 2004; Drapun & Smith 2012) by a greater number of paired spines (about 70–75) on seta-e of the first antenna in the male (in the literature 60–71 spines; probably specimens with 38–42 pairs of spines reported in M.V. Angel’s private message relating to Atlantic and Banda Sea specimens belong to another species) and seta-like filaments (about 42) (in Poulsen 1973 about 30) on the masticatory pad of the mandible.
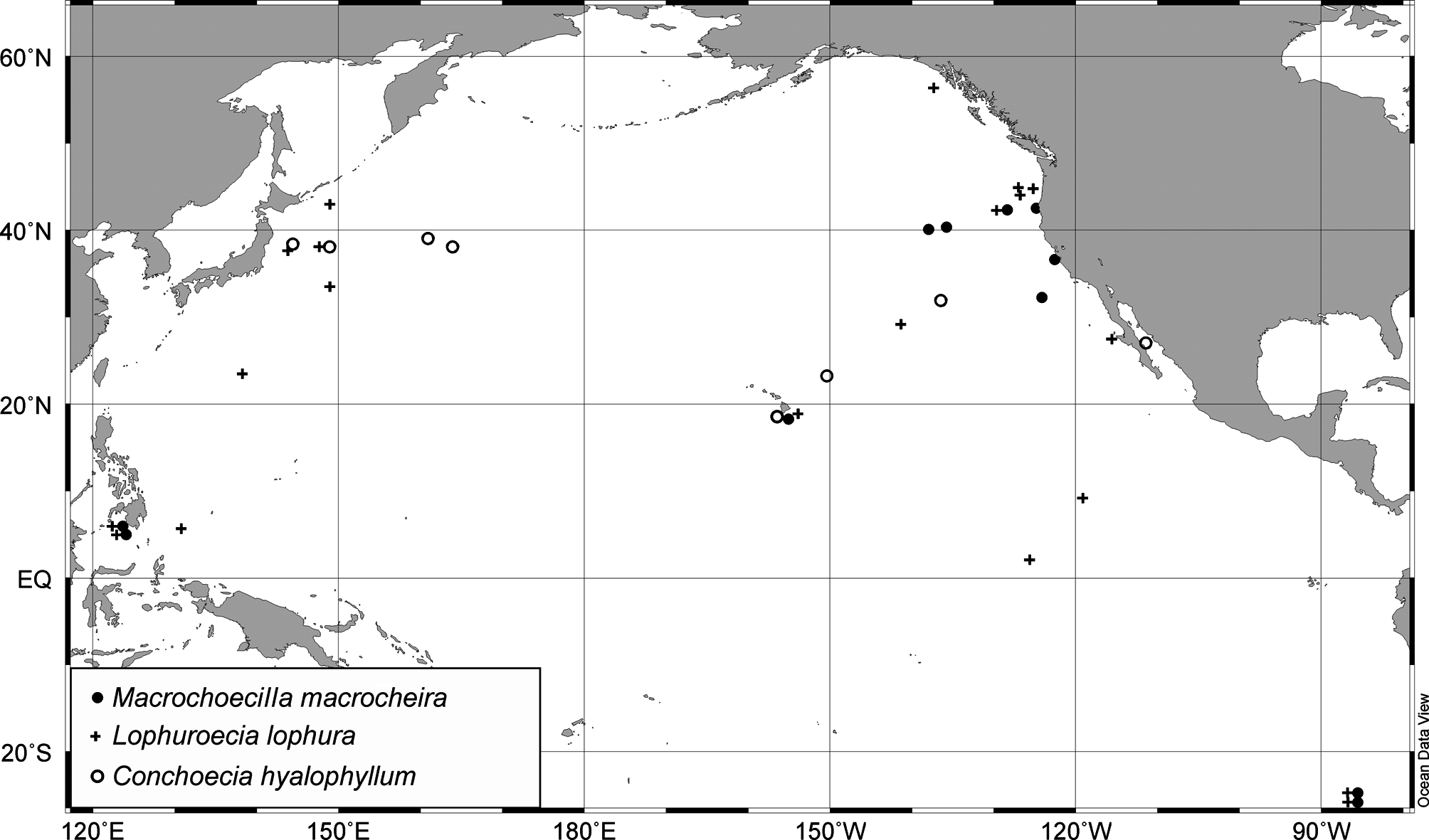
Distribution. This species has been reported from all oceans (geographical range is 60°N to 52°S), in most?temperate, subtropical and tropical zones, but with highest abundance in the tropics ( Poulsen 1973), whereas M.V. Angel (private message) reported most abundance in an area 40°–60°N in the north-east Atlantic. Its depth range is from 200–300 m to 3000 m. It is a mesopelagic species, depth range of adult males and females is usually 400– 1000 m, juveniles occur at 400– 600 m. In our material the species was collected from ( Fig. 28 View FIGURE 28 ): in the northwestern Pacific—between 5°–42°N at depths from 200–300 m to 2000–3000 m, most abundant at 600–1000 m; in the north-eastern Pacific—between 2°–56°N at depths from 200–300 m to 100–1500 m, most abundant at 500– 100 m. Hartmann & Petersen (1978) designated a syntype of this species taken at Valdivia Station 182 V 10 ˚ 08’S 97˚14’E, so the type locality is in the southern Indian Ocean.
| RV |
Collection of Leptospira Strains |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Halocypridina |
|
Family |
|
|
SubFamily |
Conchoeciinae |
|
Tribe |
Conchoeciini |
|
Genus |
Lophuroecia lophura ( Müller, 1906 )
| Chavtur, Vladimir G. & Bashmanov, Alexander G. 2018 |
Conchoecia lophura
| Drapun, I. & Smith, S. L. 2012: 54 |
Conchoecia lophura
| Angel, M. V. 1993: 176 |
| Deevey, G. B. & Brooks, A. L. 1980: 107 |
Conchoecia lophura
| Martens, J. M. 1979: 323 |
Conchoecia lophura
| Poulsen, E. M. 1973: 125 |