
Macrodactylus ocreatus Bates, 1887
|
publication ID |
https://doi.org/ 10.1649/0010-065X-64.3.193.4 |
|
persistent identifier |
https://treatment.plazi.org/id/2D50594D-AD07-D942-F9F5-FDE68ED4FA75 |
|
treatment provided by |
Diego |
|
scientific name |
Macrodactylus ocreatus Bates, 1887 |
| status |
|
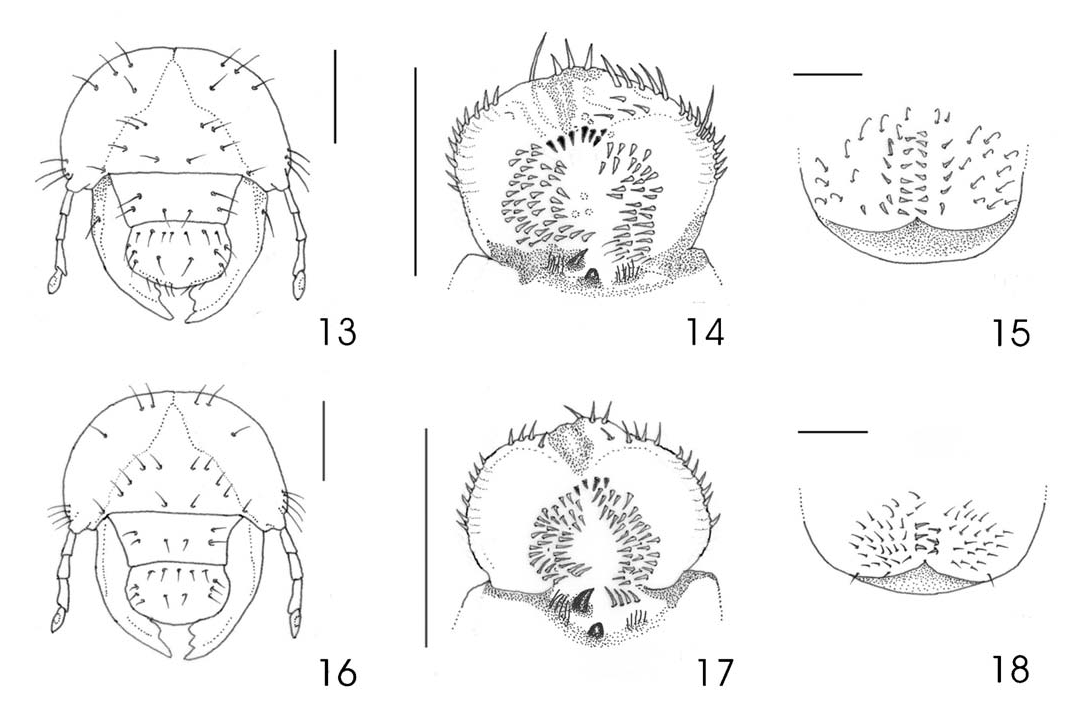
Macrodactylus ocreatus Bates, 1887 ( Figs. 16–18 View Figs )
Description is based on eight third instars collected in Mexico: Puebla, Zapotitlán Salinas, San Pedro Atzumba (18°09′41″N, 97°31′18″W), 2,000 m, 26-I-2007, from soil in amaranthus field, A. Aragón and Y. Rodríguez (DAGAM, IEXA).
Description. Head. Maximum head capsule width 2.3–2.5 mm, approximate dorsal body length 12–19 mm. Surface of cranium nearly smooth, light yellow. Frons ( Fig. 16 View Figs ) on each side with 2 exterior frontal seta, 2 posterior frontal setae, 1 anterior frontal setae, and 1 anterior angle frontal seta; remaining cranial surface with 4 dorsoepicranial setae, and 3–6 paraocellar setae on each side. Clypeus trapezoidal, with 2 lateral setae on each side and 1 anterior setae. Labrum sligthly asymmetrical, rounded at sides, with 4–5 posterior setae, 2 central setae, and 3 lateral setae on each side. Epipharynx ( Fig. 17 View Figs ) with dark, wide elongate epizygum; haptomerum with 4 short stout heli; each acanthoparia with 7 spine-like setae; acroparia with 7 long setae; corypha with 5 setae; each plegmatium with 9–10 short plegmata; both chaetoparia with many long slender setae and sensilla; pedium narrowed; dexiophoba abundant; laeophoba with many filaments; dexiotorma and laeotorma short; pternotorma rounded, narrowed; sclerotized plate large, curved; sense cone short, rounded; crepis weakly indicated and haptolachus with few crepidal setae. Scissorial area of each mandible with 2 scissorial teeth S1 and S2 fused, scissorial notch and S3 clearly defined; molar area with 2 lobes best defined in left mandible; mandibular stridulatory area absent. Mala with apical uncus of galea short, and 3 conical unci fused at their bases at apex of lacinia; maxillary stridulatory area with 10–12 small, nearly rounded teeth. Hypopharyngeal sclerome asymmetrical, produced on right side into a strong process. Last antennal segment with 1 ovate sensory spot on dorsal surface, 2 spots on ventral surface.
Thorax. Pronotum and mesonotum with transverse rows of scattered slender setae, but without spine-like setae. Thoracic spiracles light yellow, 0.33 mm long and 0.23 mm wide; lobes of respiratory plate curved, widely separated, directed posteriorly; spiracular bulla small, ovate, weakly convex. Tarsal claws similar in fore and middle legs, long with acute apex, each bearing 1 internal long seta and 1 external long seta; tarsal claw in hind legs slightly shorter than preceding, with acute apex.
Abdomen. Dorsa of segments I-VI with mixture of few rows of long setae and many spine-like setae; dorsa of segments VII-X without spine-like setae, but each with 20 long and short slender setae. Venter of abdominal segments I-IX each with a transverse row of 7–8 long setae. Respiratory plates of segments I-VIII progressively shorter (0.04–0.26 mm long; 0.09–0.18 mm wide); lobes of respiratory plates separated, directed anteriorly. Spiracular bulla slightly convex, weakly prominent. Raster ( Fig. 18 View Figs ) with one pair of longitudinal short palidia (0.20 mm total length, 0.09 mm maximum width); each palidium with 2–4 sharply pointed short pali; tegilla formed by 24–28 stout hamate setae; campus with 4 stout, long setae; barbula formed by moderate number of slender setae. Lower anal lip deeply cleft on midline; anal slit Y-shaped.
Remarks. Larvae of M. ocreatus are similar in some structures with M. mexicanus . Shared characters are number of anterior and posterior frontal setae and number of heli on epipharynx. However, M. ocreatus is easily distinguished from M. mexicanus by the presence of two exterior setae on each side of the frons, acanthoparia with seven spine-like setae, and each palidia formed only by 2–4 pali.
Distribution and Habits. Macrodactylus ocreatus is recorded from localities in México, Oaxaca, and Puebla at elevations from 1,500 to 1,700 m. Males and females are rarely found on flowers of Mimosa L. ( Mimosaceae ) (Arce-Pérez and Morón 2000).
Males and females are active from mid-April to late June. Activity is diurnal and takes place on pepper trees that are flowering but fruitless; the insects feed on pollen or fly short distances from one inflorescence to another, preferring foliage in sunny areas and remaining on branches well into the night with no apparent activity even when it is windy. Other individuals were found feeding on “huizache” flowers ( Acacia farnesiana L., Mimosaceae ). For the present study, M. ocreatus adults were found feeding on S. molle and A. farnesiana in the Valley of Tehuacán at altitudes of 1,999 and 2,253 m.
When the female is feeding on flowers, the male arrives and mounts her by grasping her with his prothoracic tarsi at the articulation of the females’ s pro- and mesothorax. With the mesothoracic tarsi, he remains attached to nearby leaves, maintaining the metathoracic legs suspended in the air, per- pendicular to the female’ s body, or supporting his body on nearby foliage. Copulation may begin immediately after mounting or within 5–20 minutes. It begins when the male’ s aedeagus is introduced into the female’ s genital organ. She continues eating during the entire process, moving from flower to flower; the male stays in the position described above. The way the bodies are oriented depends on the female’ s position before copulation–generally hanging from flowers with the ventral region facing up, so that the male hangs below the female.
Shortly before copulation ends, a brief ascendingdescending vibratory movement can be observed in the male’ s meso- and metathoracic tarsi. When the male removes his aedeagus, copulation ends and one of two things happens: a) the female starts moving, walking until the male dismounts; she continues eating and, after a time, copulates with another male; or b) the male and female stay in the mounted position, sometimes resuming copulation after a few minutes. Copulation does not occur in all such cases, as the female sometimes impedes it despite the fact that the male’ s aedeagus is exposed.
Other males frequently approach the copulating couple, sometimes causing it to relocate or to adopt a defensive action by vibrating and pushing the visiting male away with their hind legs, with both the male and female participating. On three occasions, we observed an interruption in copulation in order for the female to excrete, although we did not collect the excreta to ascertain whether or not this was a sperm selection mechanism.
Thirty-six copulations were observed, 21 in captivity and 15 in the field. In captivity, copulation averaged 80.7 min, with a minimum of 23 min and a maximum of 183 min. In the field, average duration was 49.6 min, with a minimum of 24 min and a maximum of 79 min. Copulation was sometimes interrupted after less than eight min; these events were not included in the average, as Eberhard (1993) studied the relationship between internal genital mechanisms and the outward conduct of three Macrodactylus species and concluded that brief copulations represent a failure in sperm transmission.
Our observations of the reproductive behavior of M. ocreatus coincide with those of Arce-Pérez and Morón (2000) based on studies of M. mexicanus . Adults are active in late spring and throughout summer. They copulate on the foliage of the plant they are feeding on, especially on leaves during the sunny hours. Unlike the male, the female feeds during copulation, which varies greatly in duration.
According to Eberhard (1993), it is common among species of Macrodactylus for the male to mount the female and grasp her with his front tarsi. Mounting may last just a few minutes or days. Just as variable is the time during which the aedeagus is inside the female’ s genital organ, from seconds to about 120 min for certain species. The same author observed that most of the courtship ritual occurs during and after aedeagus intromission, as the male is attempting to induce a female response that will permit the spermatophore to reach the spermatheca even after it has entered the genital chamber. This would explain why the most active movements of the male’ s meso- and metathoracic tarsi in M. ocreatus occur during the final phase of copulation, and why the male remains in a mounted position for quite a while afterwards. According to Eberhard (1893), the male may be able to cause the female to empty, at least partially, the spermatheca in order to receive the new sperm. On the other hand, M. ocreatus females can prevent copulation by walking quickly, thereby disallowing the aedeagus from entering the genital chamber; she can also kick with her back legs, as was observed by Eberhard (1993) in other species. The female may copulate several times before burying herself in the soil for oviposition, an activity that she can carry out more than once, remaining underground for hours or days.
Nineteen pairs were isolated. Six died almost immediately after capture. For the 13 surviving females, a fecundity rate of 5– 25 eggs per female was observed, with an average of 9 eggs deposited during 5 ovipositions. Carrillo and Gibson (1960) observed that M. mexicanus females deposited clusters of 5– 30 eggs in moist soil shaded by host plants, at a depth of 15–30 cm.
KEY TO KNOWN THIRD INSTARS OF MACRODACTYLUS IN MEXICO
1. Frons with 1 external frontal setae on each side. Acanthoparia formed by 9–10 spine-like setae. Each palidium with 5–8 pali........................... 2
1′. Frons with 2 external frontal setae on each side. Acanthoparia formed by 7 spine-like setae. Each palidium with 2–4 pali................. M. ocreatus
2. Epipharynx with transverse row of 4–5 heli. Each palidium with 5–6 pali...... M. mexicanus
2′. Epipharynx with transverse row of 6–7 heli. Each palidium with 6–8 pali ........ M. nigripes
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.