
Metopolophium taimyricum, Stekolshchikov & Khruleva, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4748.1.5 |
|
publication LSID |
lsid:zoobank.org:pub:A35BC3DE-1114-4731-AF3E-61956E9939DC |
|
DOI |
https://doi.org/10.5281/zenodo.3704351 |
|
persistent identifier |
https://treatment.plazi.org/id/03F90349-0873-FFBA-FF57-FD8DFB3EDE0E |
|
treatment provided by |
Plazi |
|
scientific name |
Metopolophium taimyricum |
| status |
sp. nov. |
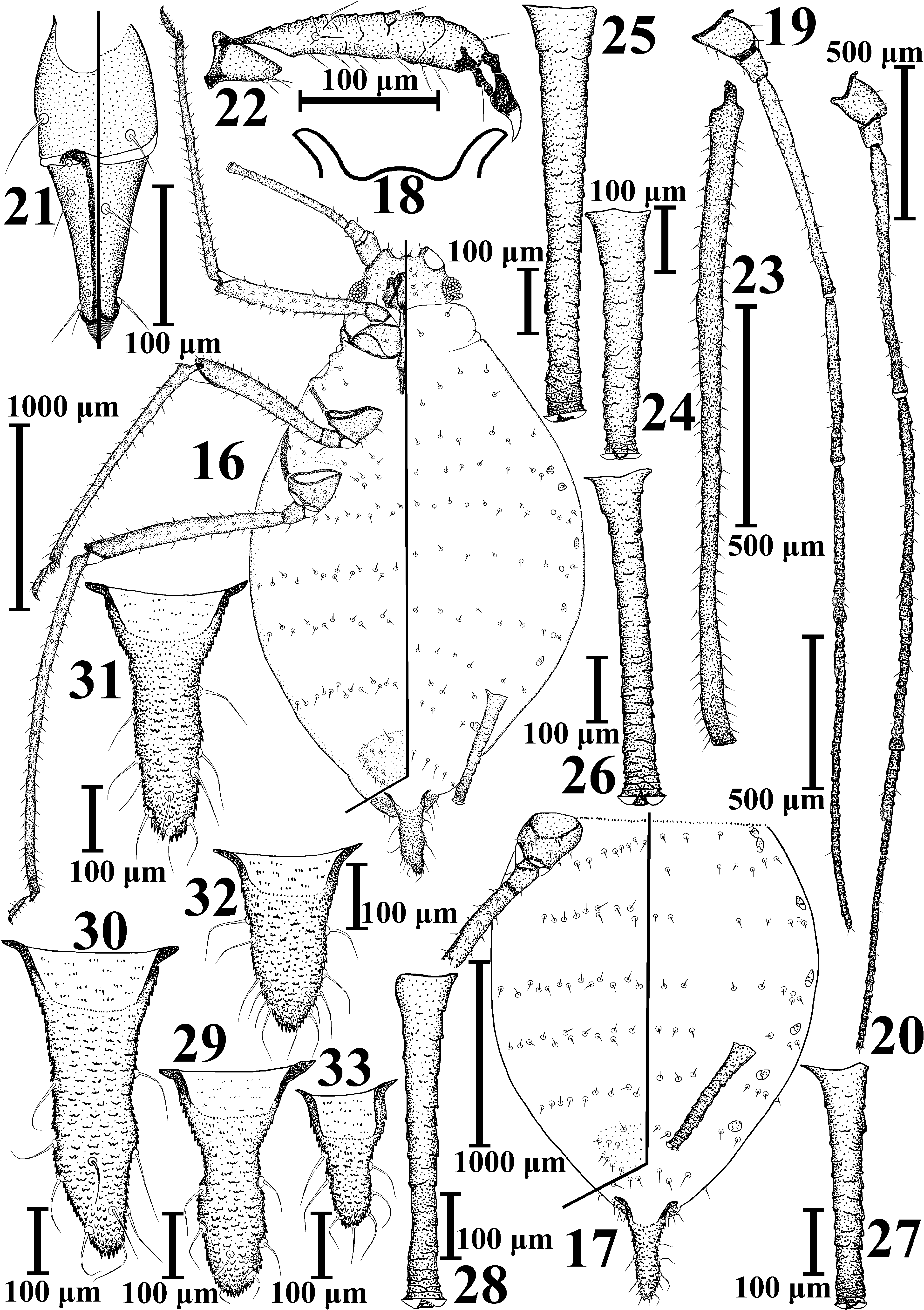
Metopolophium taimyricum sp. nov.
( Figs. 16–33 View FIGURES 16–33 , Tabl. 3–4 View TABLE 3 View TABLE 4 )
Type material. Holotype: apterous viviparous female, No. 10875, slide No. 1, specimen 3 (the left specimen in the bottom row) “ Metopolophium taimyricum sp. nov., sweeping on Hedysarum hedysaroides subsp. arcticum , Krasnoyarsk Krai, Taymyrsky Dolgano-Nenets District, near to Pyasino Lake (70° 04ˊ N, 87° 39ˊ E), 24.vii.2001, leg. О.А. Khruleva”. Paratypes: 1 fund., No. 10869, Krasnoyarsk Krai, Taymyrsky Dolgano-Nenets District, the upper course of the Nizhnaya Agapa River (70° 05ˊ N, 87° 26ˊ E), 3.vii.2001, forb-grassy meadow in a creek valley, sweeping on Hedysarum hedysaroides subsp. arcticum ; 3 apt., No. 10873, the same locality, 19.vii.2001, patchy herb-moss tundra, sweeping; 4 apt., 1 al., No. 10875, the same locality and date as holotype, the rocky top of hill with patchy legume-herb-dryad-moss tundra, sweeping on Hedysarum hedysaroides subsp. arcticum ; 4 apt., No. 10879, the same locality as No. 10869, 28.vii.2001, willow-herb-dryad-moss tundra on the top of hill, sweeping; 1 al., No. 10880, the same locality as No. 10869, 28.vii.2001, forb-grass meadow in a creek valley, sweeping; 2 apt., 1 al., No. 10883, the same locality as No. 10869, 29.vii.2001, sweeping; 4 fund., No. 10889, Krasnoyarsk Krai, Taymyrsky Dolgano-Nenets District, near to Medusa Bay, Varavikova River (13 km south of Dikson Vill., 73° 24ˊ N, 80° 44ˊ E), 22–27.vii.2002, gravelly south-facing slope of the river terrace with spotted willow-dryad-moss cover, pitfall traps; 16 fund., 2 apt., 1 al., 22 males, 6 ovip., No. 10891, the same locality as No. 10889, 2.vii–4.viii.2004, pitfall traps; 1 fund., No. 10892, Krasnoyarsk Krai, Taymyrsky Dolgano-Nenets District, near to Efremov Kamen’ Bay, Maksimovka River (32 km south of Dikson Vill., 73° 14ˊ N, 80° 44ˊ E), 5–30.vii.2004, river floodplain with willow-legume-moss cover, pitfall traps; 1 fund., No. 10893, the same locality as No. 10892, 5–30.vii.2004, gravelly dry plot in the river valley with a forb-moss-dryad cover, pitfall traps; 1 al., No. 10894, the same locality as No. 10892, 5–30.vii.2004, damp bank of river with a forb-dwarf willow-moss cover, pitfall traps; 1 fund., 1 apt., 1 ovip., No. 10908, Krasnoyarsk Krai, Taymyrsky Dolgano-Nenets District, the Ary-Mas area (72° 28ˊ N, 101° 55ˊ E), 8.vii.2010, mesophytic forb-grassy meadow with willows shrubs on the high layer floodplain, sweeping; 7 fund., 1 apt., No. 10909, the same locality as No. 10908, 11.vii.2010, dry edge of high layer of floodplain with forb-grass-dryad cover, sweeping; 1 fund., 4 apt., 1 al., No. 10911, the same locality as No. 10908, 14.vii.2010, dry sandy floodplain with forb-grass rarefied cover, sweeping; 6 apt., 1 al., No. 10914, the same locality as No. 10908, 19.vii.2010, edge of high layer floodplain with willow-herb-dryad cover, sweeping; 1 fund., No. 10915, the same locality as No. 10908, 21.vii.2010, sweeping; 1 fund., 22 apt., 3 al., 16 ovip., No. 10919, the same locality and biotope as No. 10909, 23.vii.2010, sweeping; 1 apt., 1 al., 1 ovip., No. 10921, the same locality and biotope as No. 10911, 24.vii.2010, sweeping.
Holotype is deposited at ZIN RAS; paratypes are deposited at ZIN RAS, the Natural History Museum ( London , United Kingdom) and Muséum national d’Histoire naturelle (Paris, France).
Etymology. The name taimyricum is supposed Latin demonym of Taymyr.
Description. Fundatrix. Body broadly elliptical, 1.6–1.9 (1.7) times as long as wide. Colour of living specimens unknown. Paired or unpaired marginal tubercles sometimes present on meso- and metathorax, total number of marginal tubercles on whole body 0–11 (1.8). Ratio of depth of sinus to distance between bases of antennae 0.09–0.25 (0.15). Third antennal segment with 0–4 (1.3) rhinaria near base. Ultimate rostral segment wedge-shaped or elongate wedge-shaped, 1.4–2.0 (1.7) times as long as its basal width. Peritremes on abdominal segments I and II widely spaced. Cauda finger-shaped or elongate triangular.
Apterous viviparous female. Body elongate elliptical, almost spindle-shaped, wide in the middle of abdomen (III–IV segments) and gradually getting narrow to the head and to the end of the abdomen, 1.8–2.1 (1.9) times as long as wide. Colour of living specimens unknown. Cleared specimens with antennae gradually darkening to the apex from very light-brown 1st segment to brown processus terminalis; with brown ultimate rostral segment; apices of tibiae and tarsi and the very apices of siphunculi brown; outer surface of femora light-brown; head, rostrum (except ultimate rostral segment), legs, peritremes, siphunculi, anal and subgenital plates and cauda very light-brown. Thorax and abdomen not sclerotized, pale, membranous, only VIII abdominal tergite with very light-brown and almost invisible band. Surface of head, thoracic dorsum and abdominal tergites I–VII smooth, wrinkled; cuticle on head, pro- and mesothorax intensely rugose, on metathorax and abdominal tergites I–VII weakly wrinkled, on VIII abdominal tergite with short rows of small pointed spinules, which partially fuse to form scales; thoracic venter smooth with short scales formed by small pointed spinules in base of coxae; ventral abdomen with long rows of small spinules, sometimes forming strongly elongate cells. Setae on thoracic and abdominal dorsum strongly blunt, almost rod-like, or weakly capitate; ventral thoracic and abdominal setae pointed or, rarely, blunt. Paired or unpaired marginal tubercles often present on prothorax and abdominal segments II–IV and rarely also on abdominal segment V; total number of marginal tubercles on whole body 0–10 (4.9). Marginal tubercles not large (medial diameter of siphunculus 2.3–7.0 (3.9) times as long as width of the largest marginal tubercle), from semicircular to flat. Spinal tubercles absent. Head with epicranial suture more or less distinct. Frontal tubercles well developed, antennal tubercles rather high, divergent, median tubercle not high, but clearly visible; frons with sinus, ratio of depth of sinus to distance between bases of antennae 0.16–0.34 (0.22). Setae on head blunt or weakly capitate. Antennae 6- segmented, 1st and 2nd antennal segments slightly wrinkled, almost smooth, 2nd segment ventrally with rare short scales, 3rd–6th segments with large scales which are almost invisible on 3rd segment and well-defined on 5–6th segments. Third antennal segment with 2–10 (4.0–7.0) rhinaria near base of segment. Rhinaria are round or oval, relatively small (with internal diameter 8–25 μm), very weakly protuberant, barely or not at all protruding above the surface of the segment. Antennae with blunt or weakly capitate setae on 1st–4th segments and pointed ones on 5th– 6th segments. Rostrum reaching to mesothorax. Ultimate rostral segment elongate, wedge-shaped, with slightly, but clearly convex sides, 1.7–2.2 (1.9) times as long as its basal width. Legs long, setae on legs blunt or pointed, on tarsi only pointed. Chaetotaxy of first tarsal segments 3, 3, 3 (rarely on one hind leg the first tarsal segment has 2 setae). Arms of mesosternal furca connected by wide base. Spiracles reniform. Peritremes on abdominal segments I and II separated by a distance lesser than diameter of peritreme, sometimes continuous, rarely fused or widely spaced. Siphunculi long, with distinct flange, with imbrication increasing distally, with conical base, then almost cylindrical, sometimes very slightly, almost unnoticeably swollen before flange. Subgenital plate oval, with pointed setae on anterior half and blunt or pointed setae along the hind margin. Setae on anal plate finely pointed. Cauda finger-shaped with rounded apex, often with a slight constriction at mid-length and long, finely pointed setae.
Measurements of holotype. Body—3309×1756; antenna—2925: III—718×42 (in the middle), IV—511, V— 483, VI—195+745; hind femur—1015; hind tibia—1888; siphunculus—620×61 (in the middle); cauda—455×253 (at base) ×202 (before base). For more biometric data see Tables 3–4 View TABLE 3 View TABLE 4 .
Alate viviparous female. Body elongate oval, 2.2–2.7 (2.4) times as long as wide. Colour of living specimens unknown. Cleared specimens with brown thorax. Abdomen with more or less hardly visible large light brown or brown marginal sclerites. Surface of head, pro- and mesothorax weakly wrinkled, smooth. Total number of marginal tubercles on whole body 1–8 (5.6). Head without epicranial suture. Median tubercle weakly developed. Ratio of depth of sinus to distance between bases of antennae 0.12–0.26. Third antennal segment with 7–13 (9.5) rhinaria spaced evenly along the segment, 4th and 5th segments without rhinaria. Rhinaria round or oval, sometimes relatively large (up to 30 μm), weakly protuberant, with external diameter 4.0–8.2 times as long as their height, spaced evenly along the segment. Ultimate rostral segment 1.8–2.5 (2.1) times as long as its basal width. Peritremes on abdominal segments I and II separated by a distance lesser than diameter of peritreme, sometimes continuous or fused. Cauda finger-shaped or elongate triangular with almost pointed apex.
Male. Apterous. Body elongate elliptical, almost spindle-shaped, 2.0–2.6 (2.3) times as long as wide. Colour of living specimens unknown. Abdominal dorsum sometimes more or less visible light brown, sclerotized bands on abdominal tergites VII–VIII and sometimes with spinal sclerites of different size and brown intersegmental muscle sclerites on tergites II–VI; spinal sclerites rarely fused, forming short and interrupted band. Surface of head weakly wrinkled; abdominal tergite VI with sparse, short rows of small, pointed spinules which on tergites VII–VIII partially fuse to form short scales. Paired or unpaired marginal tubercles sometimes present on prothorax and on abdominal segments I–IV; total number of marginal tubercles on whole body 0–5 (1.3). Head without epicranial suture. Median tubercle low, inconspicuous, ratio of depth of sinus to distance between bases of antennae 0.09–0.21 (0.16). Third antennal segment with 26–52 (35.5), 4th segment with 17–33 (25.9), 5th antennal segment with 13–27 (18.1), and base of 6th antennal segment with 0–1 (0.1) rhinaria. Rostrum reaching to meso- or metathorax. Peri- tremes on abdominal segments I and II separated by a distance lesser than diameter of peritreme. Cauda elongate triangular with almost pointed apex.
Oviparous female. Body elliptical, 1.8–2.1 (1.9) times as long as wide. Colour of living specimens unknown. Total number of marginal tubercles on whole body 0–8 (2.2). Median tubercle weakly developed. Third antennal segment with 0–3 (0.5–1.5) secondary rhinaria. Peritremes on abdominal segments I and II separated by a distance lesser than diameter of peritreme. Hind tibiae clearly swollen in basal half, with 75–156 round or oval small pheromone plates located in basal two thirds of tibia.
Distribution. Known only from Krasnoyarsk Krai, Taymyrsky Dolgano-Nenets District.
Biology. Life cycle unknown. The simultaneous presence of the fundatrices, apterous and alatae viviparous and oviparous females in the same sample indicates that the life cycle of the species is short. The first fundatrices were noted at the beginning of July and an oviparous female was found in one of the samples (No. 10908), together with them, on July 8. The rapid appearance of the ovipara could only happen if she was born directly of the fundatrix. The presence in sample No. 10919 of July 23, of a large number of apterous parthenogenetic and oviparous females together with a living fundatrix, can also be explained by the latter’s ability to produce both of these morphs. Otherwise, it is difficult to imagine how in northern conditions parthenogenetic females, born in early July, could complete development and give birth to oviparous females by the time of the collection. There is no doubt that the life cycle of the new species consists of 2–3, or at most 4 generations, which makes it impossible for it to be a dioecious species, involving migration from a primary to a secondary host. That is, this species is monoecious.
The distribution of M. taimyricum is interesting in that it is quite widespread in the tundra zone of Taymyr, but absent outside of it (including in the forest-tundra transition zone). In the arctic tundra, the new species was collected together with M. arcticum sp. nov. in a single biotope with early melting snow and one of the warmest for this area. In typical tundra, M. taimyricum was found in three biotopes located in the river valley, about 1 km from each other. In the southern tundras of Taymyr, M. taimyricum was found in various habitats, often with Acyrthosiphon pisum . The largest series of the new species was collected in dry biotopes with legume-herb-dryad cover ( Fig. 34 View FIGURE 34 ).
Most species of Metopolophium , for which the host plants are known, develop on Rosaceae and Poaceae , but four have been described from plants of other families, including two species described from the Canadian Arctic: M. arctogenicolens Richards from Taraxacum arctogenum Dahlstedt (Asteraceae) and M. pedicularis (Richards) from Pedicularis sudetica Willd. ( Orobanchaceae ). In the southern tundra, M. taimyricum was repeatedly collected in sweep samples on the common Hedysarum hedysaroides subsp. arcticum (Fabaceae) . This plant is also noted, but with a small abundance, from north-western Taymyr ( Matveyeva & Zanokha 1997). It is possible that it may be a host plant for M. taimyricum although the true host plant of this species, as that of M. arcticum , has not been established definitively.
Systematic relationships. The new species is placed in the genus Metopolophium , and neither Acyrthosiphon nor the Sitobion subgenus Metobion , on the basis of the same characters as M. arcticum sp. nov.: (1) the presence of a well-developed median frontal tubercle ( Fig. 18 View FIGURES 16–33 ) and (2) the rostrum reaching the mesothorax with its ultimate segment more than 0.7 times longer than the second segment of hind tarsus.
As mentioned above, 21 species are currently included in the genus Metopolophium ( Favret 2019) . The differences between apterous viviparous females of all these species (with the exception of M. arctogenicolens whose apterous viviparous females are unknown) and those of Metopolophium taimyricum are detailed in Table 5 View TABLE 5 . Unfortunately, based on the characteristics in the table, the new species cannot be distinguished from M. pedicularis . Metopolophium taimyricum is also very similar to M. darjeelingense , M. dirhodum , M. fasciatum and M. festucae .
Metopolophium pedicularis was described by Richards (1972) from four apterous viviparous females, one alatae viviparous female, 12 oviparous females and two males. The description was very short including a small number of morphological characters. Metopolophium pedicularis differs from the new species in having relatively short setae: Richards (1972) indicates that for apterous viviparous females of M. pedicularis , the setae on tergite VIII are shorter than half the diameter of the siphunculi as measured immediately basal to flange (0.74–1.21 for M. taimyricum ). The alatae viviparous female M. pedicularis has more rhinaria on the 3rd antennal segment– –17 (7–13 for M. taimyricum ), more setae on the ultimate segment of rostrum– –9 (6–7 for M. taimyricum ), and fewer setae on the cauda– –7 (9–12 for M. taimyricum ), his PT/BASE ratio is greater– –4.00 (2.57–3.76 for M. taimyricum ), and the URS/HTII ratio is less– –0.67 (0.75–0.87 for M. taimyricum ). The oviparous females of M. pedicularis have a slightly higher PT/BASE ratio—3.57–4.71 (2.48–3.63 for M. taimyricum ) and sometimes more rhinaria on the 3rd antennal segment– –2–5 (0–3 for M. taimyricum ). Siphunculi of males of M. pedicularis are significantly longer (450–550) than those of males of M. taimyricum (261–354).
The new species also differs from M. darjeelingense in that the latter has 4 setae on the first segment of the hind tarsi, whereas M. taimyricum has 3. An additional difference with M. dirhodum is that the antennal segments 3–6 of apterous viviparous females of M. dirhodum are pale with dusky apices, while the antennal flagellum of apterous M. taimyricum is gradually darker from base to apex. The function “length of processus terminalis × length of siphunculi”/“length of cauda × length of 2nd joint of hind tibia” is 6.00–8.60 for apterae of M. fasciatum and 3.89–6.37 for apterae of M. taimyricum , dorsal abdominal setae on tergite III of apterae of M. fasciatum maximally 10–16 long (17–33 for M. taimyricum ) and spinal tubercles usually present at least on one side of occiput of apterae of M. fasciatum (completely absent in M. taimyricum ). The apterous viviparous female of M. festucae has a relatively short ultimate rostral segment– –89–119 (114–142 for M. taimyricum ), and the head and abdominal tergites VII and VIII of apterous of M. festucae are frequently with spinal tubercles (always absent in M. taimyricum ). Metopolophium taimyricum differs from a number of species ( M. albidum , M. alpinum , M. chandrani , M. dirhodum , M. fasciatum , M. festucae , M. lacheni , M. montanum and M. tenerum ) by the presence of apterous males, whereas in these species the males are alatae. Apterous viviparous females of Metopolophium arctogenicolens are unknown, but the oviparous female differs from the oviparous female of M. taimyricum in the ratio of the length of the ultimate rostral segment and the length of the 2nd segment of the hind tarsus (0.71 for M. arctogenicolens and 0.77–0.91 for M. taimyricum ), a large number of additional setae on the ultimate rostral segment (8–11 for M. arctogenicolens and 4–6 for M. taimyricum ), and a large number of rhinaria on the 3rd antennal segment (3–8 for M. arctogenicolens and 0–3 for M. taimyricum ). Males of M. arctogenicolens differ from males of M. taimyricum by the ratio of the length of the ultimate rostral segment and the length of the 2nd segment of the hind tarsus (0.71 for M. arctogenicolens and 0.76–0.91 for M. taimyricum ), a large number of additional setae on the ultimate rostral segment (8 for M. arctogenicolens and 4–6 for M. taimyricum ), and by the ratio of the length of the processus terminalis and the length of base of the 6th antennal segment (3.33 for M. arctogenicolens and 3.48–5.37 for M. taimyricum ).
From the second new species described in this article, M. arcticum sp. nov., M. taimyricum sp. nov. differs by the fact that in M. taimyricum oviparous females URS/HT2 is 0.77–0.91 (0.94–1.07 in M. arcticum ), PT/BASE is 2.48–3.63 (1.56–2.35 in M. arcticum ), as well as many other features, including characters of fundatrices and males (see Tables 1 View TABLE 1 , 3 View TABLE 3 , 4 View TABLE 4 ).
The new species differs by the following characters from the apterous viviparous females of twelve Acyrthosiphon species inhabiting the north and listed at the section “Systematic relationships “ of the description of M. arcticum sp. nov.: A. auctum , A. boreale , A. brevicorne and A. malvae have URS/HT2 more than 0.90 (0.73–0.89 in M. taimyricum ); A. assiniboinensis , A. ignotum , A. knechteli and A. pisum have ratio of length of siphunculi and length of body more than 0.22 (0.15–0.22 in M. taimyricum ); A. svalbardicum has PT/BASE 1.18–1.52 (2.31–3.92 in M. taimyricum ); A. loti and A. brachysiphon have very short setae that on the 3rd antennal segment are shorter than 0.50 the articular diameter of the segment and on abdominal tergite VIII shorter than 0.80 of this diameter (0.47–0.86 and 1.07–1.90 in M. taimyricum , respectively); A. loti also has small number of secondary rhinaria on 3rd antennal segment of apterae– –0–3 (2–10 in M. taimyricum ); A. brachysiphon has a ratio of length of antenna and length of body 0.90–1.30 (0.74–1.01 in M. taimyricum ), and PT/BASE is 3.30–4.80 (2.31–3.92 in M. taimyricum ). Only A. churchillense presents some problem for comparison with the new species because it is very briefly described and its description does not include many of the necessary characters. At the same time, from the description of A. churchillense it is clear that specimens of all morphs have very short setae on 3rd antennal segment– –10 and a relatively short ultimate rostral segment– –100–110––with 2–6 accessory setae (15–25, 109–142 and 4–8 in M. taimyricum , respectively).
Pleotrichophorus glandulosus (Kaltenbach)
Material. NDU: 8.vii.2017, Artemisia vulgaris , shaking, fund. ( AS) .
Published information. NSV: ( Ivanovskaya 1977).
Uroleucon (Uromelan) aeneum (Hille Ris Lambers)
Material. NUO: near to Talnakh, 26.vii.2017, Cirsium helenioides , on stem, apt. ( AS) .
Uroleucon (Uromelan) campanulae (Kaltenbach)
Material. NDU: 3.vii.2017, Campanula rotundifolia L., on apices of stem, fund. ( AS) . NUO: near to Talnakh, 24–25.vii.2017, Campanula rotundifolia L., on apices of stem near flowers, apt., al. ( AS) .
Uroleucon (Uromelan) jaceae jaceae (Linnaeus)
Published information. NSV: ( Ivanovskaya 1977).
Material. NUO: near to Talnakh, 24.vii.1964, Asteraceae , fund . ( EI).
Uroleucon (Uromelan) solidaginis (Fabricius)
Material. NUO: near to Talnakh, 6.viii.1964, Asteraceae or Alnus sp., apt. and al. ( AE); 6.viii.1964, Alnus sp., fund. ( EI); 25.vii.2017, Solidago virgaurea subsp. dahurica (Kitag.) Kitag. , on flower stalk, apt., al. ( AS).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Aphidinea |
|
Family |
|
|
SubFamily |
Aphidinae |
|
Tribe |
Macrosiphini |
|
SubTribe |
Macrosiphina |
|
Genus |