
Mideopsis milankovici, Pešić & Smit, 2020
|
publication ID |
https://doi.org/ 10.24349/acarologia/20204387 |
|
publication LSID |
lsid:zoobank.org:pub:93EBC5FF-0F17-41BA-9BBB-5EE840E8A84C |
|
persistent identifier |
https://treatment.plazi.org/id/B619AD87-81F8-4156-B86E-3360053AF3F8 |
|
taxon LSID |
lsid:zoobank.org:act:B619AD87-81F8-4156-B86E-3360053AF3F8 |
|
treatment provided by |
Marcus |
|
scientific name |
Mideopsis milankovici |
| status |
sp. nov. |
Mideopsis milankovici sp. nov.
Zoobank: B619 AD 87-81F8-4156-B86E-3360053AF3F8
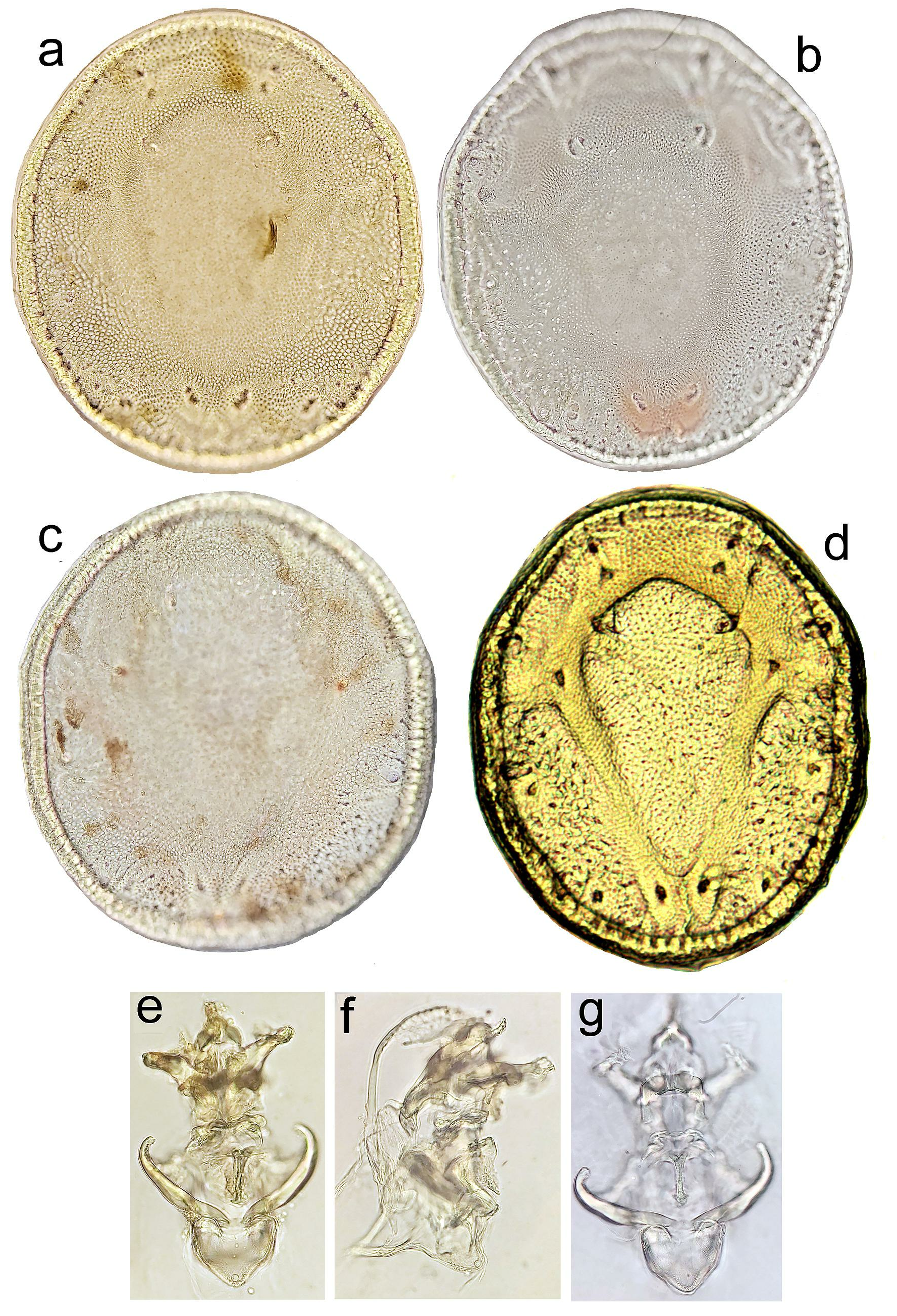
Figures 2–3 View Figure 2 View Figure 3 , 4a–b, e–g View Figure 4
Material examined — Holotype ♂ ( RMNH), sequenced [22. M19_24_2_E12], dissected and slide mounted, Montenegro, Bar , Međurječka rijeka stream, downstream, between villages of Pečurice and Krute, 42°01 ′ 21.21 ″ N, 19°13 ′ 11.92 ″ E, 26.vi.2019, leg. Pešić. GoogleMaps Paratype: 5♂♂, 17♀♀, Bar, Međurječka rijeka stream, upstream, 42°2 ′ 10.80 ″ N, 19°13 ′ 4.30 ″ E, 16.vi.2020,
leg. Pešić, 1♂, 1♀ dissected and slide mounted ( RMNH).
Other material — Montenegro, Bar, Stari Bar old town, Rikavac steam, 42°6 ′ 1.72 ″ N, 19°8 ′ 33.03 ″ E, 3.vi.2021, leg. Pešić 1♀.
Diagnosis — Dorsal shield flattened, in the centre slightly elevated, V-shaped area formed by anteriorly diverging lines of fine porosity little evident. Ejaculatory complex with well sclerotized anterior keel, anterior ramus wedge-shaped. Postgenital area short (males 116–125 µm, 18% dorsal shield L; females 85 µm, 12% dorsal shield L), excretory pore closer to posterior idiosoma margin (distance 25–35 µm).
Description — Idiosoma rounded; colour dark yellowish to brown. Dorsal shield in the and within (diagonal) studied mideopsid species.
centre slightly elevated, with anteriorly diverging lines of particularly fine porosity forming a V-shaped area sligthly evident ( Figures 4a–c View Figure 4 ). Postgenital area short (about 12–18% dorsal shield L), excretory pore close to posterior idiosoma margin (distance 25–35 µm). Ejaculatory complex with a strongly sclerotized anterior keel, anterior ramus wedge-shaped ( Figures 2c–d View Figure 2 ). Palp: P-1 with a seta in it’s proximal part; P-2 ventral margin almost straight; P-3 ventral margin concave, distal margin convex; P-4 ventral projection directed ventrally, with two tips, both flanked by a fine subterminal seta; distal part of P-4 slightly narrower than basal part, with 3 subapical setae, one ventrally, slightly thicker and stiff, and two fine, located dorsally and laterally ( Figures 2e View Figure 2 and 3b View Figure 3 ). Male – P-2 L/H ratio 2.5. Female – P-2 more slender than in male, L/H ratio 2.7.
Measurements — Male (Holotype; in parentheses paratype, n = 1) – Dorsal shield L
684 (641), W 606 (575), L/W ratio 1.13 (1.12). Ventral shield L 800 (772), W 800 (747); gnathosomal bay L 149 (141), Cx-III W 416 (378), distance between IV-L insertions 466 (419). Genital field: gonopore L/W 153/50 (156/50), ratio 3.1 (3.1), L Ac-1-3: 44 (41), 44 (45), 44
(41). Distance genital field-excretory pore 77 (75), excretory pore-caudal idiosoma margin 34
(30). Ejaculatory complex L 244 (275). Capitulum vL 130; chelicera: total L (163), claw L
(92). Palp: total L 240 (227), dL/H, dL/H ratio: P-1, 28/31, 0.92 (30/30, 1.0); P-2, 67/45, 1.48
(59/42, 1.41); P-3, 31/37, 0.84 (31/34, 0.93); P-4, 73/29, 2.5 (69/28, 2.46); P-5, 41/16, 2.65
(38/16, 2.4); L ratio P-2/P-4 0.93 (0.86). dL of I-L-1-6: 61 (61), 63 (68), 65 (62), 77 (67), 103
(98), 127 (128); I-L-6 H 36 (36); dL/H I-L-6 ratio 3.5 (3.6). dL of IV-L-1-6: 84 (77), 106 (91),
84 (78), 113 (116), 138 (132), 131 (134).
Female (paratype, n = 1) – Dorsal shield L 708, W 625, L/W ratio 1.13. Ventral shield L 750,
W 775; gnathosomal bay L 153, Cx-III W 391, distance between IV-L insertions 447. Genital field: gonopore L/W 141/86, ratio 1.6; L Ac-1-3: 47, 47, 38. Distance genital field-excretory pore 44, excretory pore-caudal idiosoma margin 25. Chelicera total L 164, claw L 84. Palp: total L 242, dL/H, dL/H ratio: P-1, 34/30, 1.16; P-2, 59/44, 1.36; P-3, 31/36, 0.88; P-4, 77/28,
2.73; P-5, 41/15, 2.65; L ratio P-2/P-4 0.78. dL of I-L-1-6: 52, 61, 63, 70, 97, 122; dL of
IV-L-1-6: 77, 106, 82, 125, 141, 138.
Etymology — Named after Prof Milutin Milanković (1879–1958), the eminent Serbian astrophysicist best known for developing one of the most significant theories relating to earth movements and long-term climate change.
Discussion — The phylogenetic analysis based on COI data reveals that M. milankovici sp. nov. is most similar to M. orbicularis . The high distance between these two species (18.8% K2P) suggests a long independent history of these two species. The relatively high K2P distance seems to be typical for water mites ( Blattner et al. 2019) and the obtained data of our study are comparable with the genetic distance between cryptic species of other water mite clades (see Stålstedt et al. 2013, Martin et al. 2010, Pešić et al. 2017, 2019a). Mideopsis orbicularis is widely distributed in the Palaearctic, inhabiting various types of standing waters such as lakes and canals, occasionally also lowland streams ( Gerecke et al. 2016), but never recorded from sites with a seasonal flow. From a morphological point of view, M. orbicularis can be separated from the new species only in the male sex, based on the shape of the ejaculatory complex (anterior ramus regularly rounded, anterior keel narrow and weakly sclerotized – see Figure 3c View Figure 3 ). Mideopsis roztoczensis , a species widely distributed in running waters in Europe,
is characterized by a more elevated dorsal shield, a larger postgenital area (140–190 µm), the excretory pore more distanced from the posterior margin of the idiosoma (distance in general 40–90 µm), and the anterior ramus of the male ejaculatory complex (wedge-shaped as in M. milankovici sp. nov.) being wider, with a characteristic arrow-shaped delimited area – see Figure 3d View Figure 3 ).
Due to similarity in shape of ejaculatory complexes (see Figure 3e View Figure 3 ), M. milankovici sp.
nov. resembles M. persicus Pešić & Saboori, 2015 . The latter species is known from a single male collected in a stream in Fars Province of South Iran ( Pešić and Saboori 2015). Mideopsis persicus can be separated by the shape of the dorsal shield with distinct, anteriorly diverging lines of fine porosity forming a well visible V-shaped area. In the new species from Montenengro this V-shaped area is almost indistinguishable (compare Figures 4a–c View Figure 4 with Figure 4d View Figure 4 ).
Mideopsis crassipes , a species widely distributed in the Holarctic ( Gerecke et al. 2016)
and M. rossicus , a species known from Russia ( Tuzovskij 2002), can be separated from all above-mentioned species including M. milankovici sp. nov. in having an egg-shaped idiosoma and a ventral extension of P-4 strongly curved with anteriorly directed tips.
Habitat — Characteristics of sampling sites indicate a preference for intermittent habitats. Both streams in which M. milankovici sp. nov. was collected are located in the narrow coastal region of Montenegro, their middle and lower courses regularly dry up in summer (for an overview of the species and communities that inhabit intermittent rivers in the southern part of Montenegro see Pešić et al. 2020). The upper part of the Međurječka rijeka stream is perennial ( Figure 5 View Figure 5 ) but runs dry in its lower reach.
Distribution — Montenegro.



| RMNH |
National Museum of Natural History, Naturalis |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |