
Muscari erzincanicum Eker, 2021
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.487.1.3 |
|
persistent identifier |
https://treatment.plazi.org/id/03BFFC30-9D0C-FFF5-B88F-C3D931C1FC0C |
|
treatment provided by |
Marcus |
|
scientific name |
Muscari erzincanicum Eker |
| status |
sp. nov. |
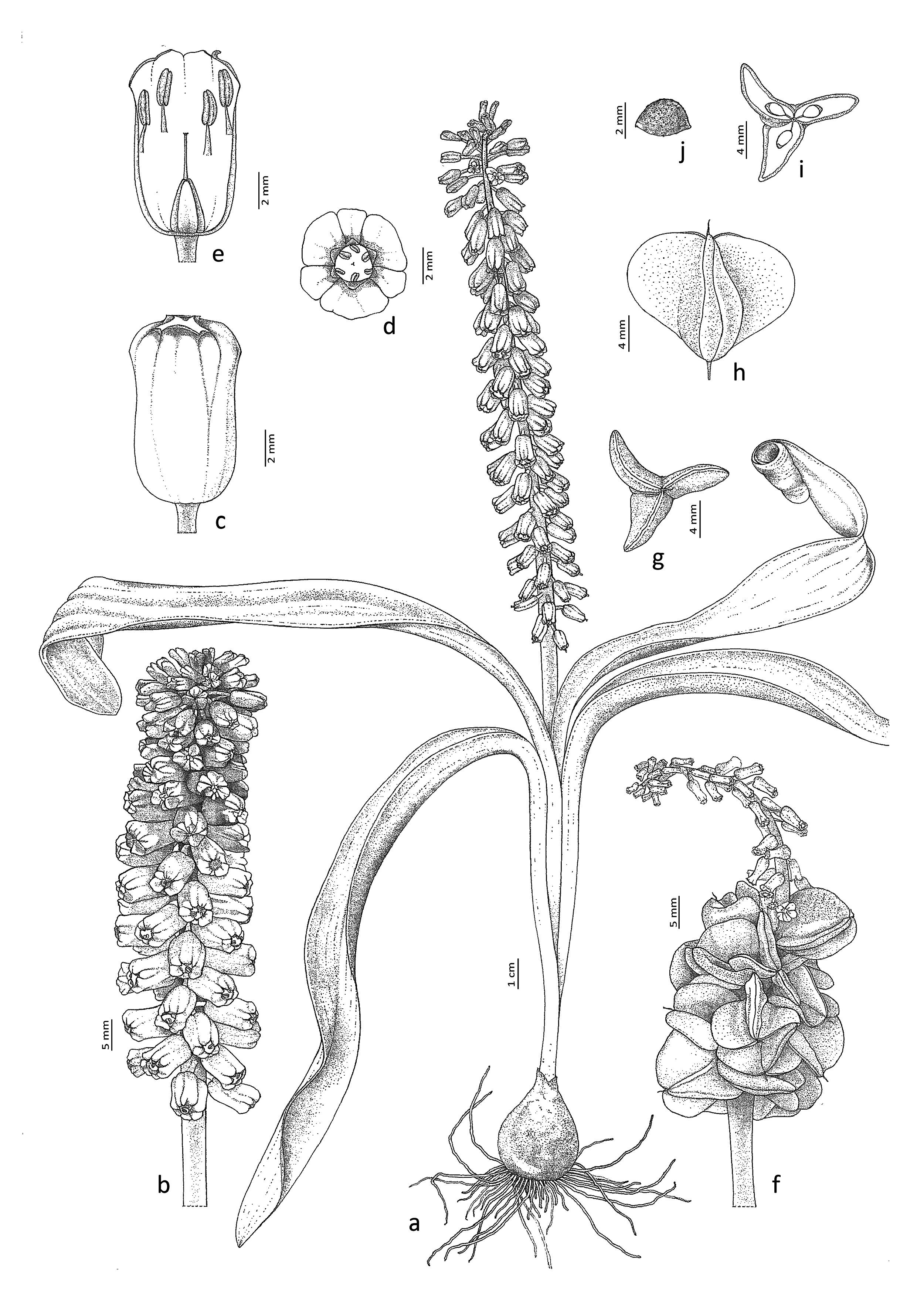
Muscari erzincanicum Eker View in CoL sp. nov. ( Figs. 2–5 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 ).
Muscari erzincanicum is similar to M. massayanum and M. erdalii , but is easily distinguished from M. massayanum and M. erdalii by its distinctly emarginate and broadly oblate capsule with wavy valves, not elongating pedicel of fertil flowers in fruit and dull black colored seeds. From M. massayanum it also differs by the characteristics of outer tunics, leaf number per scape, cylindrical and more compact raceme in flower and fruit, oblong and wider fertile flowers, wider orifice, color of young intermediate flowers, seed shape, seed ornamentation, testa epidermal cells shape, anticlinal cell walls. From M. erdalii it also differs by the ovoid bulb, leaf number per scape, glabrous and wider leaves, dense to moderately dense and narrow raceme in flower, shorter pedicel of fertil flowers in flower, color of young intermediate flowers, type of cuticle on the seed body, anticlinal and periclinal cell walls.
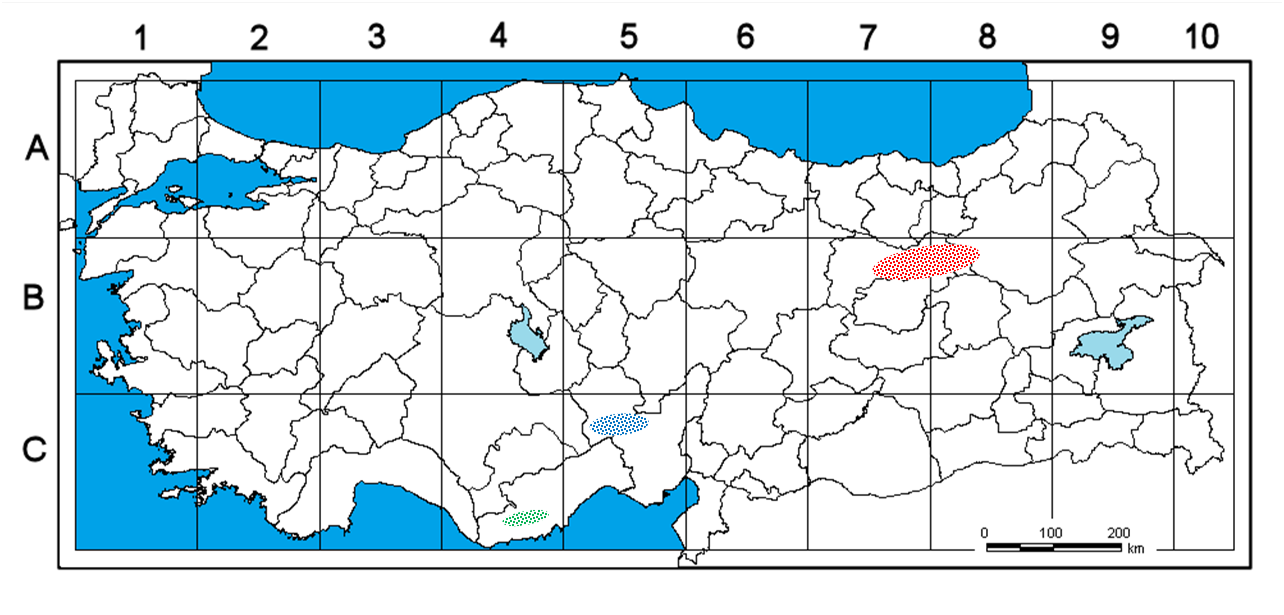
Type:— TURKEY. B7 Erzincan: Between Erzincan to Refahiye, surroundings of Sakaltutan Pass , on fine serpentine stacks, 1531 m of elevation, 08 May 2020 , İ. Eker 12931 , A. Kandemir , E. Eker (holotype AIBU!; isotypes AIBU!, HUB!) .
Paratypes:— TURKEY. B7 Erzincan: between Erzincan-Refahiye, surroundings of Sakaltutan Pass , on fine serpentine stacks, 1531 m of elevation, 12 June 2020 , İ. Eker 12968 (in fruit) ( AIBU!); ibid., slope, 2600 m of elevation, 11 July 2005 , M. Koyuncu 15353 & E. Özhatay (in fruit) ( AEF 21931!); ibid., Sakaltutan Mountain , stony slopes, 2000 m of elevation, 18 June 2005 , M. Koyuncu 14996 & N. Aslan ( AEF 24008!); ibid., Sakaltutan Mountain , slope, 2450 m of elevation , M. Koyuncu 15106 & N. Aslan ( AEF 24256!); Sakaltutan Mountain Pass , while coming to Erzincan Plain, stony slope, 1950 m , M. Koyuncu 15018 & N. Aslan ( AEF 24081!); ibid., 19 June 2005 , M. Koyuncu 15011 & N. Aslan (in fruit) ( AEF 24088!); between Balkaya-Karaören, slopes, 1725 m of elevation, 28 May 2005 , M. Koyuncu 14887 & N. Özhatay ( AEF 24153!); Büyükköşkünler ( Başpınar ), hills, steppe, 1400–1450 m of elevation, 05 July 1967 , İ. Döl (in fruit) ( ISTF 22380 About ISTF !); Kemah-Tanközü ( Tan Village ), 1250 m of elevation, 02 June 1975 , T. Baytop s.n. (in fruit) ( ISTE 31877 About ISTE !); Keşiş Mountain, above Yaylakent Village , steppe slope, 2300−2500 m of elevation, 13 July 2005 , M. Koyuncu 15387 & N. Özhatay (in fruit) ( AEF 24972!); Yaylabaşı Town , gravelly slopes, 1400 m of elevation, 27 May 1998 , A. A. Dönmez 6104 ( HUB!); Tunceli: North of Pülümür , c. 2000 m of elevation , E. Pasche 78/18 (cult. E 00345640 !) ; B8 Erzurum: hills WNW of Aşkale , loose igneous scree on steppe, c. 2000 m of elevation, 17 July 1986 , J. C. Archibad 7815 (in fruit) ( E 00129956 !) .
Bulb ovoid, 2.5−5 × 2.5−4.5 cm, without bulblets. Outer tunics papery, pale brown; inner tunics membranous, creamy and often white dotted. Leaves 1–4 per scape, mostly humistrate or sometimes patent, recurved at tips, broadly linear to linear-lanceolate and attenuate at base, thick, deeply canaliculate, 15−45 × 1−3 cm, sinuate at margin, glabrous, acute and cucullate. Scape 1 per bulb, stout, erect to flexuose, 9–25 cm long, shorter than leaves, not elongating in fruit. Raceme dense to moderately dense, cylindrical, 5−15 × 1.5−2 cm in flower, not elongating in fruit and fruiting raceme remaining dense and compact. Pedicels of fertile flowers 0.5 – 2 mm long, shorter than perianth, patent to slightly deflexed, not elongating in fruit. Fertile flowers oblong, 7−11 × 4−6 mm, strongly shouldered and strongly constricted distally (orifice 1.5–2 mm), greenish or pinkish-green in bud, young intermediate flowers purple or pinkish-purple with greenish or greyish-green tips and shoulders, and turn to greenish-yellow or ivory at maturity; lobes c. 0.5 mm long, blackish, slightly recurved. Pedicels of sterile flowers 1–7 mm long, semierect to patent, bluish-violet colored, mostly shorter than sterile flowers or sometimes longer. Sterile flowers obconical to cylindrical-obovoid, 3.5–6.5 × 1.5–3 mm, shorter than fertile ones, bluish-violet colored with pinkish tips. Stamens biseriate, filaments 1–1.5 mm long, whitish, upper series attached about 3 mm below the mouth of tube and lower series 1 mm below upper ones, anthers purplish-black, 1.5–2 mm long, pollen light yellow. Ovary green, ovoid, 2–3.5 × 1.5–2.5 mm; style whitish, 2–3 mm long. Capsule broadly oblate, distinctly emarginate, with strongly compressed wing-like and wavy valves, 12–16 × 15–22 mm. Seeds 3−3.5 × 2.5−3 mm, broadly elliptic-ovoid; surface reticulate to rugose, black. Flowering and fruiting in May–June ( Table 1).
Etymology: —The specific epithet of new species is derived from the province of Erzincan, where the new species was first discovered and is more abundant than other regions.
The Turkish name for the new species: — Muscari is called “ Müşkürüm ” or “ Sümbül ” in Turkish. The author proposes “ Erzincan sümbülü ” as a vernacular name for M. erzincanicum according to the guidelines of Menemen et al. (2016).
Distribution and habitat: — Muscari erzincanicum has a wide range of natural distribution especially in the Erzincan surroundings, Pülümür/ Tunceli and Aşkale/ Erzurum ( Fig. 7 View FIGURE 7 ). The distribution of the plant extends to Kemaliye district in the south, to north of Pülümür in the southeast direction, to Aşkale in the northeast, and is interrupted by the Kelkit Valley and the mountain ranges separating the Black Sea and inner-eastern Anatolia in the north. The region is very rich in terms of endemic species. Muscari erzincanicum grows on fine serpentine stacks, stony and gravelly slopes, and loose igneous scree on steppe at 1250 – 2600 m of elevation. The new species is a plant belonging to the Irano-Turanian floristic region.
Species conservation assessment: —As a result of combining of the herbarium records and the locations determined in field studies on the map, it is understood that M. erzincanicum spreads over an area (EOO) of approximately 10.000 km 2. It can be predicted that a large number of populations and/or locations of the new species can be determined in addition to these records. The habitats in the areas where the new species spreads are generally natural areas away from settlements and are not under obvious threats. Therefore, the species may be classified as ‘Least Concern’ (LC) ( IUCN 2019).
Comparative seed micromorphology with close Muscari species :— Muscari erzincanicum differs from M. massayanum in terms of seed shape in outline (broadly elliptic-ovoid vs. ovoid to globose), seed color (dull black vs shiny black), sculpture ornamentation (reticulate to rugose vs. pustulate and pustulate-foveate), testa epidermal cells shape (polygonal, oblong vs orbicular to elliptic), and anticlinal cell walls (regularly thickened and raised to straight, partly undulate vs sunken, ribbed, undulate). Muscari erzincanicum also differs from M. erdalii in terms of seed color (dull black vs shiny black), type of cuticle on the seed body (rugose vs smooth), anticlinal cell walls (regularly thickened and raised to straight, partly undulate vs irregularly thickened, minutely raised to straight), and periclinal cell walls (convex or flat vs concave to flat) ( Table 2, Fig 6 View FIGURE 6 ).
Taxonomic relationships and Discussion: —When the records of Muscari massayanum in the Flora of Turkey ( Davis & Stuart 1984) are combined with the other herbarium specimens examined, it appears on the map that two groups of populations are aggregated on the southwestern and northeastern ends of the Anatolian Diagonal. A third group of populations (Mut/ Mersin) is assembled beyond the southwestern end of the diagonal ( Fig. 7 View FIGURE 7 ). Although the Anatolian Diagonal emerged as a barrier separating Anatolia into two, during the last glacial period (115000–12000 years ago) it also served as a bridge between the North and the South, and moreover between the Caucasus and the Mediterranean vegetation. During plant migration, new species emerged, which were probably unable to move north again during warmer periods and whose genetic link was cut off ( Davis, 1971, Ekim & Güner 1986, Gür 2017). Here, the northern and southern populations, which probably had the same origin, diverged over time by the different topographic, soil and climatic characteristics they encountered. For example, a similar distribution is also in question for the Anatolian endemics Muscari azureum Fenzl (1859: 165) and Muscari coeleste Fomin (1908: 11) , which are very similar morphologically to each other. On the other hand, the herbarium sample of Huber-Morath (coll. no: 10355) which Davis & Stuart (1984) regarded as M. massayanum , could not found in herbarium collections. However, it was understood that the samples collected by the author of this study at the specified location in this record were M. erdalii . Among the close related taxa of the new species mentioned, M. erdalii is a distinct species with many distinctive morphological characters (e.g. globose bulb, distinctly scabrid leaves, longer pedicel of fertil flowers, not winged-looking fruits with slightly compressed valves) whose details are given in the “taxonomy section”. The habitat type of M. erdalii spreading over limestone bedrock is also quite different from the other two taxa. Muscari erdalii , despite being published as a new species by Demirci et al. (2013), Huber-Morath ’s record “ 10355 ” given from the same region in the Flora of Turkey was not mentioned therein and no comparison was made with its close relative M. massayanum . However herbarium specimen Cowley et al. “ 72/90-334 (1990)” collected from the same region, after Davis & Stuart (1984), was shown among the paratypes.
The southern populations, Muscari massayanum sensu Eker , are located in the Mediterranean phytogeographical region, while northern populations, M. erzincanicum , are distributed in the Irano-Turanian phytogeographical region. Also, morphologically Muscari erzincanicum differs significantly from M. massayanum especially by its cylindrical and more compact raceme in flower and fruit, oblong and wider fertile flowers, purple or pinkish-purple young intermediate flowers, distinctly emarginate bigger fruits with wavy valves, not elongating pedicels of fertil flowers in fruit, seed shape, seed color and numerous seed surface ornamentation characteristics ( Table 1–2; Figs. 1–6 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 ).
In this study, it was observed that seed surface ornamentation and some other seed characteristics in the genus Muscari can be used in species distinction. In this regard, extensive and detailed studies of seed surface properties are incomplete in the genus Muscari . It was reported that epidermal characters are less influenced by environmental conditions and they provide valuable criteria for classification at interspecific and family level with their high structural diversity ( Barthlott 1981). In addition, detailing genetic-based differences in generative structures such as seed surface, which are less affected by environmental impacts, will shed light on the taxonomy of the genus Muscari . A detailed seed micromorphology study of the taxa of Hyacinthaceae (= Asparagaceae subfamily Scilloideae ) spreading in the South Africa was conducted by Brudermann et al. (2019). A research to be carried out on the same family spreading especially in the Mediterranean basin, will probably contribute to the elucidating the systematic relationships of both the genus Muscari and the family Hyacinthaceae .
In previous studies, flowers of Muscari subgen. Leopoldia were examined in two categories as fertile and sterile ones, and included in species descriptions. Here, the fertile flowers are divided into two according to their development stage for the first time; Intermediate and mature ones. Intermediate flowers are fresh flowers in the prematurity stage that found between mature and sterile flowers and show color characteristics that can be used in species distinction in subgen. Leopoldia ( Fig. 5 View FIGURE 5 ). The mature flowers are generally not discriminative for many species in the subgen. Leopoldia, as they often turn to shades of ivory and brown colors. But young intermediate flowers have more characteristic features in color. Therefore, this character state in Leopoldia may be significant, and it should be tested whether it is distinctive among other close taxa.





| İ |
"Alexandru Ioan Cuza" University |
| A |
Harvard University - Arnold Arboretum |
| E |
Royal Botanic Garden Edinburgh |
| AIBU |
Abant Izzet Baysal Üniversitesi |
| HUB |
Hacettepe University |
| M |
Botanische Staatssammlung München |
| AEF |
University of Ankara |
| N |
Nanjing University |
| T |
Tavera, Department of Geology and Geophysics |
| J |
University of the Witwatersrand |
| C |
University of Copenhagen |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.