
Neobelba pseudopapillipes Bulanova-Zachvatkina, 1967
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3796.2.9 |
|
publication LSID |
lsid:zoobank.org:pub:267D829B-9941-4A01-BA67-AA9C877F5413 |
|
DOI |
https://doi.org/10.5281/zenodo.6133923 |
|
persistent identifier |
https://treatment.plazi.org/id/0383C80D-B631-A91E-588B-FB03FBA9E2CD |
|
treatment provided by |
Plazi |
|
scientific name |
Neobelba pseudopapillipes Bulanova-Zachvatkina, 1967 |
| status |
|
Redescription of Neobelba pseudopapillipes Bulanova-Zachvatkina, 1967 View in CoL
Diagnosis (adult, immatures unknown). Damaeid mite of medium size with increased number of setae on trochanters and femora: setal formula of trochanters 1-1-4-3, setal formula of femora 10-10-9-9. Solenidia of all legs, with exception of φ 1 on tibia I and φ on tibia IV, relatively short. All solenidia of genu I – III and tibia II – IV with associated companion seta d. Prodorsum without propodolateral apophysis P, with one pair of tubercles in dorsosejugal area (B a–B p). Both sensillus and interlamellar seta with elongated and attenuated, flagellate tips. Distinct round area (apd) resembling area porosa present anterior to insertion of interlamellar seta. Notogastral setae, at least in the posterior part of notogaster, clearly bent and characteristically thickened near base, when seen in lateral view. Ventral side without apparent tubercles. Discidium narrow, bent posterolaterad, claw-like.
Material examined. Neotype and 14 paraneotypes: Russia, Voronezh region, Ramonskiy district, forest near village of Aydarovo, 51°54’02”N, 39°18’05”E, about 157 m a.s.l.: mixed broad-leaved forest with dominance of Quercus spp ., Populus tremula , Betula pendula and Pinus silvestris , in the forest floor; collected 16.6.2012, leg. V. Kolesnikov.
The type series will be deposited in the following collections: The neotype and 2 paraneotypes in the Acarological Collections of Senckenberg Museum für Naturkunde Görlitz ( Germany), 2 paraneotypes in Arachnological Collections in zoological department of the National Museum in Prague ( Czech Republic), 3 paraneotypes in private collection of Ladislav Miko and remaining 7 paraneotypes in collection of V. Kolesnikov.
Further material examined (not included in the neotype series). Slovakia, East Slovak region, Košice – Kavečany, in the area of zoological gardens, 48°47’25”N, 21°11’54”E), 430 m a.s.l., mixed broad-leaved forest with dominance of Carpinus betulus and Quercus spp ., with deforested grassy patches, in the litter. 4 individuals, collected 2.8.1989, leg Ľubomír Kováč. 2 individuals deposited in Arachnological Collections in zoological department of the National Museum in Prague ( Czech Republic), 2 individuals in private collection of Ladislav Miko.
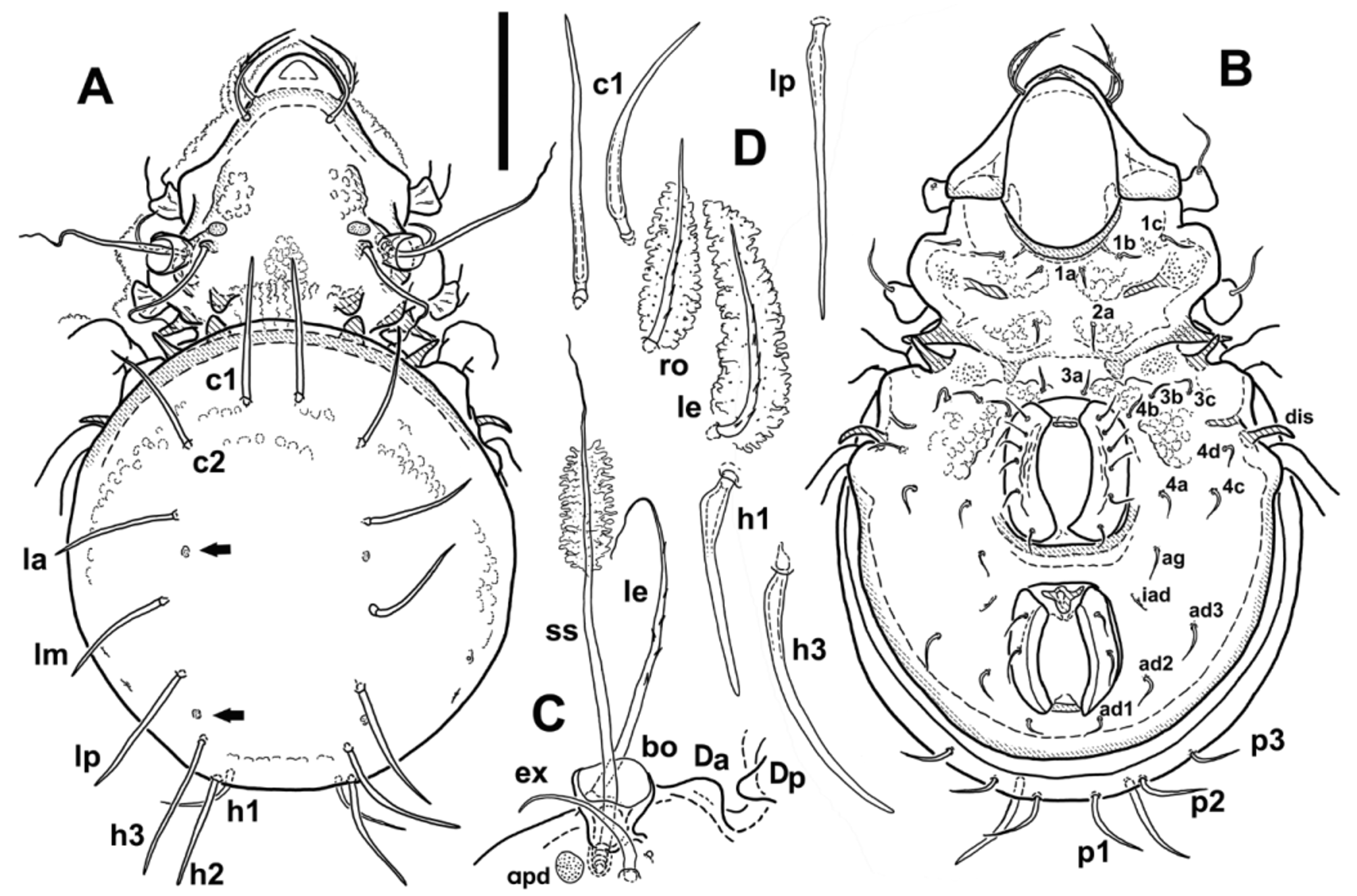
Description (adult). Figs 1 – 4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 . Dimensions Total body length 480 – 530 (510), ventral length 430 – 498 (452); maximal width of notogaster 260 – 300 (278), maximal width of prodorsum 185 – 230 (208); n=10. Length of legs and other detailed measurements in Table 1.
Lengths of setae (all in µm), n=5
Prodorsum Notogaster Ventral part of body
ro 67 (62 – 70) c1 96 (90 – 99) 1a 31 (30 – 32) le 75 (70 – 78) l a 86 (81 – 90) 2a 32 (30 – 34) in 124 (115 – 130) l p 92 (87 – 97) 3c 37 (34 – 39) ex 37 (34 – 39) h3 95 (92 – 98) 4b 38 (37 – 39) ss 165 (155 – 170) h1 64 (63 – 66) g1 28 (27 – 31) p1 60 (58 – 62) ag 37 (34 – 38) p3 34 (33 – 36) an1 34 (30 – 38) ad1 40 (38 – 41) Lengths of legs and leg segments (n=5).
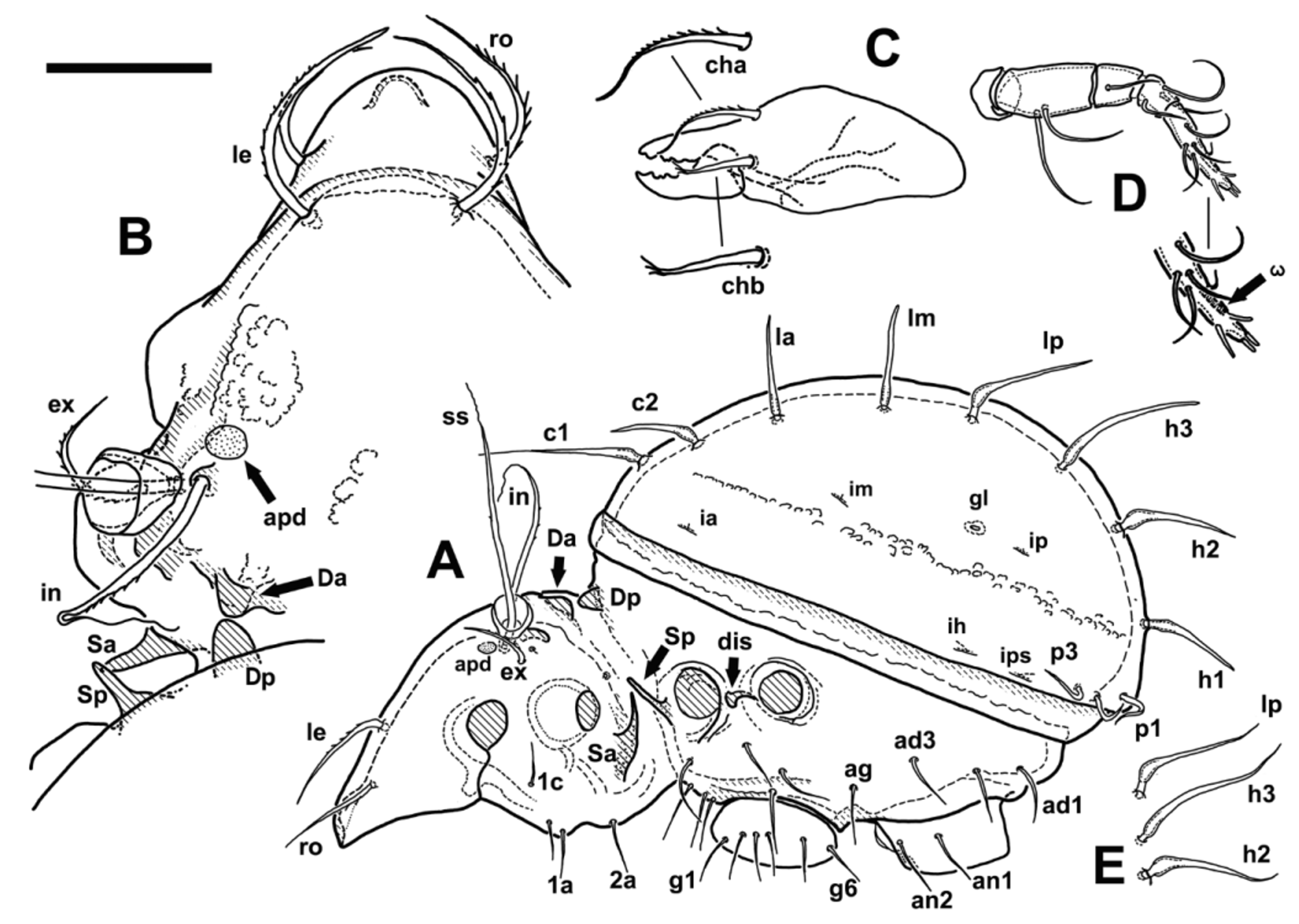
Integument. Color medium reddish brown, individuals from Slovakia lighter, yellowish brown. Almost whole body surface covered by thick layer of cerotegument, mostly amorphous and/or granular (epimeral area), with filamentous excrescences present as well, being most prominent in sejugal area. Rostral and lamellar seta covered by cerotegument, except of tip of ro ( Fig. 1 View FIGURE 1 D). Club-like bunch of filamentous and amorphous cerotegument present on distal part of sensillus, about between half and three quarters of its length, just before distal attenuated part ( Fig. 1 View FIGURE 1 C). Notogastral setae without cerotegument cover. Legs covered by thick layer of cerotegument, except distal part of all tarsi (only showed on Figs 3 View FIGURE 3 and 4 View FIGURE 4 , in other Figs omitted). Underlying cuticle smooth or microtuberculate. Adults bearing gastronotic exuviae of immature stages, often with small particles of adhered foreign material, exuviae however loosely attached and often lost during extraction.
Prodorsum. Figs 1 View FIGURE 1 A, 2A, 2B. Prodorsum conical, broadly triangular in dorsal view, sharply narrowed in sejugal area. Rostrum rounded, centrally with weakened cuticle appearing dorsally as lighter triangular area. Insertions of rostral setae (ro) on small tubercles, which are positioned at end of short ridges or lines running parallel to prodorsum edge. Insertions of lamellar setae (le) on posterior edge of arch, created by more sclerotised ribs running along lateral edge of prodorsum, and further between le insertions, in this area less visible and forming 2-3 less distinct lines. Apophysis P absent, edge of prodorsum lateral to bothridia almost rounded, in some cases most lateral part of prodorsum with short, straight edges, subparallel to body axis. Single enantiophyse present in dorsosejugal area (considered to be centrodorsal enantiophyse D, for questions of homology see discussion in Remarks), well developed, with anterior tubercle (Da) smaller, broadly triangular and sitting on sclerotised oblique ridge running from posterior end of bothridial area. Posterior centrodorsal tubercle (Dp) larger, about twice longer than Da, conical, its base hidden under anterior edge of notogaster. Enantiophyses B (postbothridial) and L (lateral) absent, but slight elevations and thickenings of cuticle above the apophyse S a and posterior to bothridia may in dorsal view resemble flat tubercles L a.and B a. Parastigmatic apophyses spiniform, oblique, with tips overlapping in dorsal view. S a with broader base, and, when seen ventrally, much longer than (about twice as long as) S p. Apophyse S p more narrow, with subparallel edges, better observable in ventral view, sharp-tiped. Bothridium positioned on elevated and more sclerotised area, short but distinct ridge runs from its base anteromediad. Interlamellar seta (in) inserted on small, distinct tubercle. Small, pore-like opening present behind insertion of ex ( Fig 1 View FIGURE 1 C, 2A), distinct pore visible also in lateral view behind acetabulum II in sejugal area ( Fig 2 View FIGURE 2 A). Unusual, distinctly pronounced area, resembling porose areas of other oribatids, present anterior to in insertion (prodorsal area, apd). Rostral (ro) and lamellar (le) setae of subequal length, curved inwards, with spinuli (le) or barbs (ro) on outer curvature, le slightly stronger at their base. Interlamellar seta (in) longer than ro and le, with sparse barbs in middle part and with long, flagellate tip, the latter often broken away ( Fig. 1 View FIGURE 1 C, 2B). Exobothridial seta (ex) relatively long, directed anteriad, bent, gradually attenuated distally, setiform. Bothridial funnel as usual in Damaeidae , cup-like, relatively large, positioned relatively far laterally. Distinct fields of sigillae (muscle insertions) present on prodorsum, two anteriad—anteriomediad to bothridia and in, one in interbothridial area. Lateral edges of two anterior fields strengthened and more sclerotised, creating a kind of narrow ridge.
Notogaster. Figs 1 View FIGURE 1 A, 2A. Notogaster circular or sub-circular in dorsal view, in lateral view almost hemispherical, very slightly eccentric with maximum height slightly posterior to center. Notogastral setae strong, setiform to spiniform, relatively long, pointing more or less radially outwards of notogaster. In dorsal view they may appear straight or almost straight ( Fig. 1 View FIGURE 1 D), but almost all clearly bent when seen in lateral view. Proximal part of some setae, seen in lateral view, with distinct and very characteristic thickening, with external edge curved proximally almost rectangularly ( Fig. 1 View FIGURE 1 D– h1, Fig. 2 View FIGURE 2 A). In some individuals, some notogastral setae (usually of rows l or h) may be bent outwards distally, creating slightly S-form shape in lateral view ( Fig. 2 View FIGURE 2 E). Posterior notogastral setae (p1-p3) shorter, bent, without thickening at base, pointing laterad or posterolaterad. Circumgastric row of muscle sigillae distinct, well visible. Two pairs of minute light spots present on notogaster posteriad or posteromediad to insertions of setae la and lp (indicated by black arrows on Fig. 1 View FIGURE 1 A).
Gnathosoma . Subcapitulum as usual for family, diarthric, with three pairs of setae. Setae fine, setiform, curved, in ventral view seta m appearing longest, but in lateral view almost equal to h, seta a slightly shorter. Setal formula of palp 0-2-1-3-9(1), solenidion ω adhered to surface of palp tarsus and hard to observe ( Fig. 2 View FIGURE 2 D). Chelicera of shape and relative size as usual in Damaeidae , seta cha longer, bent outwards proximally and inwards distally, with short barbs on external curvature and attenuated tip, seta chb straight, distally bent and with fringe of diminishing barbs ( Fig. 2 View FIGURE 2 C).
Epimeral region. Fig 1 View FIGURE 1 B. Mentotectum relatively narrow, low. Ventral tubercles absent, however, pair of triangular internal cuticular thickenings present at place of posterior ventrosejugal tubercles (Vp), which may be misinterpreted as tubercles. Ventrosejugal groove relatively broad and quite deep, laterally distinctly bordered by posterior edge of elongated apophyse S a. Epimeres III with standard set of 3 setae, epimeral setal formula 3-1-3-4. Epimeral setae subequal in length, fine, setiform and smooth, shorter than other setae of body. Epimeres with muscular sigillae, well visible, particularly closer to axial part.
Anogenital region. Fig. 1 View FIGURE 1 B. Discidium (dis) narrow, elongated and bent backwards, claw-like. Genital opening larger than anal opening, circumgenital area framed by indistinct anterior and posterior ridge, surface of genital plates along setal insertions rugged. Preanal sclerite with two distinct lateral projections. Setal formula of anogenital region as in other Damaeidae : genital g: 6; aggenital ag: 1; anal an: 2; adanal ad: 3. Adanal setae, particularly ad3, inserted far from anal opening and closer to lateral edge of ventral plate. All anogenital setae smooth, setiform, subequal in length, similar to epimeral setae. Lyrifissure iad apoanal—oblique and divergent from body axis posteriad.
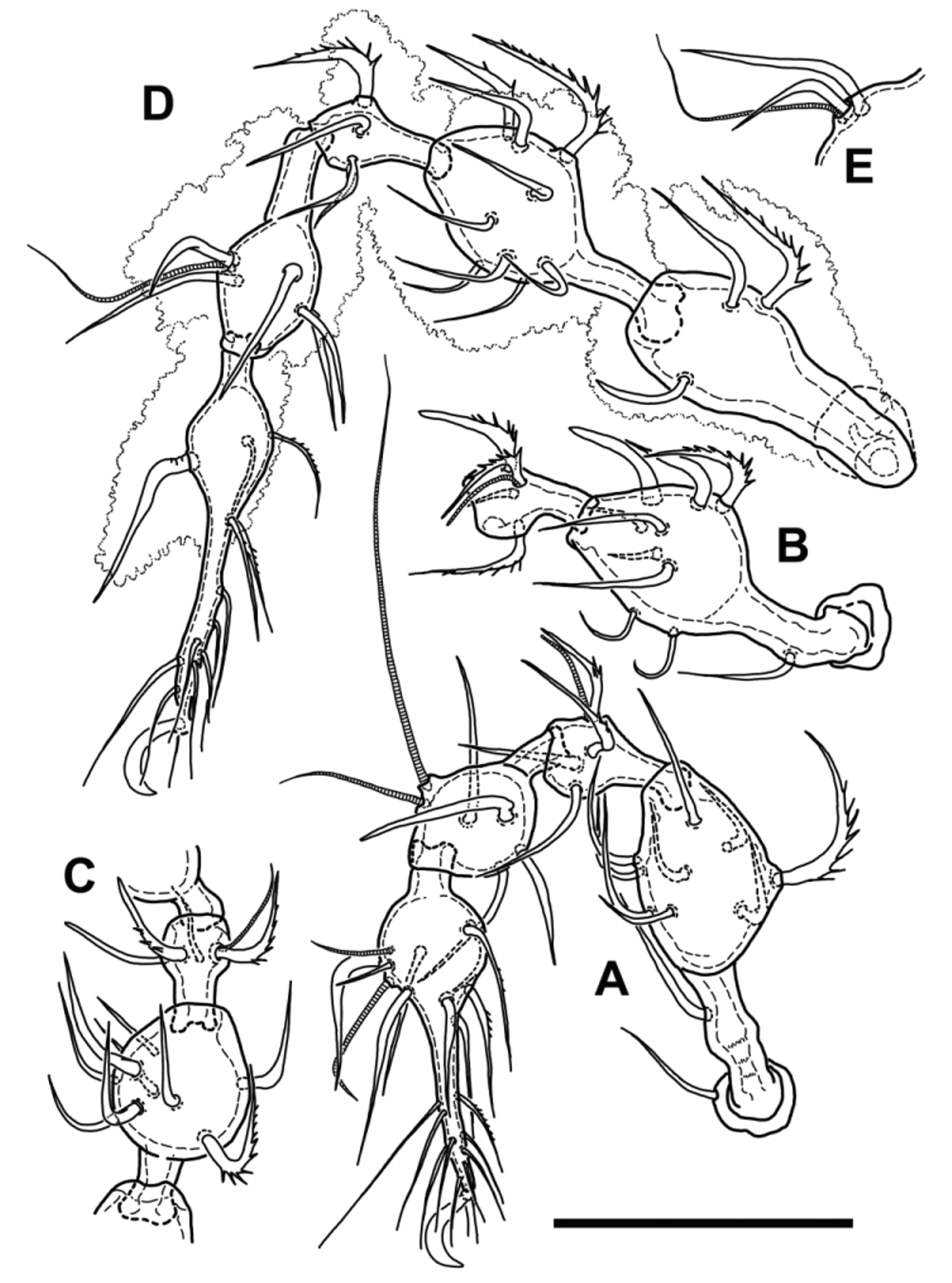
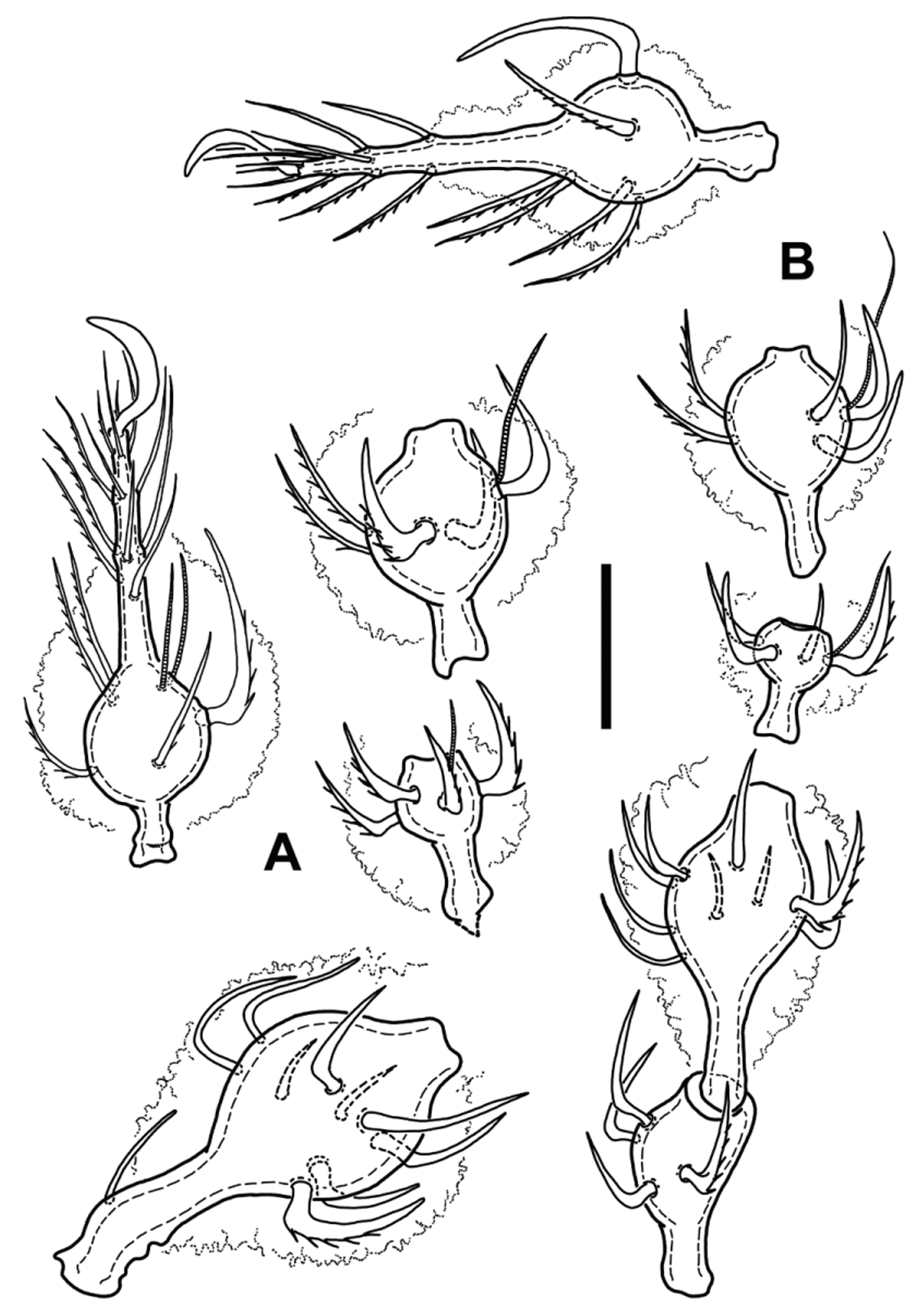
Legs. Figs 3 View FIGURE 3 , 4 View FIGURE 4 . Legs monodactylous, relatively short (leg I shorter than body, leg IV about as long as body length, slightly longer than ventral length, other legs also shorter than body, and overall length of legs increasing from leg I to leg IV—see Table 1). Leg segments with distinctly swollen distal parts (moniliform), except tarsi, where swollen part is proximal. Leg setae strong, setiform to spiniform, short to medium long, always shorter than their respective segment, mostly slightly bent, often with barbs or small spines at their outer curvatures. Dorsal setae of trochanters, femora, genua, and seta ft” of all tarsi very strong, robust, strongly and almost rectangularly bent proximally, mostly with long distinct spines at external curvature. Lateral setae of genua and tibiae usually similarly developed, but slightly smaller, other setae more narrow, or almost simply setiform, more straight and usually shorter. Leg setal formula except of seta d on tibia IV identical to species of Metabelba , as follows: leg I: 1- 10-4 [1]-4(2)-20(2); leg II: 1-10-4 [1]-5[1]-17(2); leg III: 4-9-4[1]-5[1]-17(0); leg IV: 3-9-4-5[1]-14(0). Famulus (e) normal, setiform, emergent, relatively long. Solenidia of genu I, II and III each coupled with companion seta d, as well as solenidia of tibia II, III and IV. Length of solenidia on genua subequal to length of companion seta, tibial solenidia II–III slightly longer than respective companion setae, tibial solenidion IV about twice as long as companion seta or even slightly longer ( Fig. 3 View FIGURE 3 D, E). Solenidia of tibia I free as typical for Damaeidae , setiform, tactile, φ1 very long, about 2,8 times longer than φ2. Solenidia of tarsus I relatively short, setiform, pointed, solenidion ω1 about twice as long as ω2, distally bent. Solenidia of tarsus II straight, blunt, subequal in length.
Ontogeny. Unknown.
Geographical distribution and ecology. N. pseudopapillipes is recorded only very rarely. Except classical localities of Bulanova-Zachvatkina (1967) in Moldova and Central Russia, and two localities presented here ( Slovakia and Russia—Voronezh region), we are aware of only two other potential records, both from Italy—one from Alps in province of Trentino ( Baratti et al., 2000), from ski slopes in elevation of about 1600-1700 m a.s.l., and another one from beech and oak forests in southern Tuscany, from elevation between 300 and 800 m a.s.l. ( Migliorini et al., 2002). However, in both cases the species found was reported as “ N. cf. pseudopapillipes ”, so there is no definitive clarity, if the species indeed belonged to N. pseudopapillipes . Subías (2004, 2013) reported this species from “Europa Meridional”, but it remains unclear to us on which basis. Looking on confirmed localities of the species, it may be expected in lowland, alluvial and midland forests, dominated by Quercus and Carpinus , even if it cannot be excluded that the species occurs also in other type of litter of the forests in the (?southern) temperate zone and also in forest-steppic habitats in the south-eastern Europe. The species may be therefore expected, besides known localities in Russia, Moldova and Slovakia, also in Ukraine, Kazakhstan, Romania and Hungary at least.
Remarks on species and its generic status. N. pseudopapillipes as re-described here fits very well to known characters of the species, as defined by Bulanova-Zachvatkina (1967, 1975) and reviewed above. Nevertheless, few differences appeared and raised the question of the identity of our material with original pseudopapillipes . The observed discrepancies relate to the sejugal enantiophyses. While in original description there is recorded unspecified presence of paired tubercles in dorsosejugal area, and no ventral tubercles mentioned, in updated version Bulanova-Zachvatkina (1975) states, that dorsal tubercles should be B1 and B2, i.e anterior and posterior postbothridial tubercle (B a and B p as they are named today). In the same place, the presence of posterior ventrosejugal tubercle V2 is recorded (V p in recent notation).
In present work, we consider (consistently with situation in several Metabelba species) the present dorsal tubercles to be D a and D p, based on their rather central position axial to the level of in insertions. Nevertheless, homology of these tubercles remains unclear, as they may be more linked to the development of latero – posterior rather than postero-axial end of bothridial cuticular protuberance. This is supported by the argumentation of Behan Pelletier & Norton (1985) and by the fact that protuberance on which is positioned bothridium is in N. pseudopapillipes connected with anterior tubercle of this enantiophysis by oblique ridge ( Fig. 2 View FIGURE 2 B). Shift of the whole enantiophyse in axial direction may be also explained by narrowing of the whole body in the sejugal area. Regarding presence of ventrosejugal posterior tubercle V p (corresponding to V 2 of Bulanova-Zachvatkina), we note that cuticular triangular thickenings in corresponding area may be interpreted as apophyses, when seen purely in ventral view. Nevertheless, observation from another angle (lateral) shows that there is no protuberance present, and tubercle V p is missing.
As all other characters fit exactly the description of Bulanova-Zachvatkina, and no other similar species was found yet, we consider the identity of our individuals with Neobelba pseudopapillipes proved.
The species shows important similarities with species of the genus Metabelba Grandjean, 1936 . Development of prodorsum and notogaster and their setae, characters of ventral part of the body as well as the legs are identical or very similar. As far as we know, the following characters can be considered as unique for pseudopapillipes : (a) presence of companion seta d, coupled with solenidion on tibia IV, (b) presence of distinct prodorsal area (apd), resembling area porosa, anterior to insertion of interlamellar seta in, (c) characteristic form of notogastral setae, bent and thickened at the base in lateral view. Absence or presence of accompanied setae related to tibial solenidia is broadly accepted as important distinguishing character at generic or even supra-generic level; however, this mostly applies to total absence (all tibial solenidia free) than to the absence of individual companion seta. On the other hand, setation of the leg IV seems to be of greater importance than in legs II and III, and absence of companion seta on tibia IV was used as one of the distinguishing characters, e.g., between Belba von Heyden, 1826 and Caenobelba Norton, 1979 or Dyobelba Norton, 1980 . Subías (2004) placed genus Neobelba as a subgenus to genus Metabelba , and later ( Subías, 2013) even synonymized it with Metabelba s. str. We assume that the motivation for this move was in great morphological similarity of N. pseudopapillipes and species of the genus Metabelba . Mourek et al. (2012) considered Neobelba as a separate genus, until more information about morphology of this taxon will be available, and mentioned namely the chaetotaxy of leg femora. To decide about the status of Neobelba it is crucial to weight the value of presence vs. loss of seta d on tibia IV, accompanying the respective solenidion. As demonstrated above, this character can be used as distinguishing between the genera. This supports the idea that presence or loss of seta d on tibia IV is rather a generic than a specific character within Damaeidae . In all cases where it differentiates between the genera, however, this character is combined with some additional distinguishing characters. Two other distinguishing characters of N. pseudopapillipes mentioned above can hardly be seen as generic: variability of notogastral setae within the genus Metabelba is very broad, and areas similar to prodorsal area apd appear on different places and different species throughout the family. This leaves only single generic character to distinguish Neobelba from Metabelba .
In our view, the only solution of the problem is in placing Neobelba to broader Metabelba concept at the subgeneric level. The proposed synonymy of Neobelba and Metabelba s. str. must be rejected. Nominal subgenus of Metabelba has well developed propodolateral apophysis P, which is clearly missing in Neobelba . On the other hand, N. pseudopapillipes closely resembles some of the species of recently defined subgenus Pateribelba Mourek, Miko & Bernini, 2012. It has to be noted that notation of the prodorsal tubercles in Mourek et al. (2012) followed the same approach of designation based on the physical position of tubercles (axially to the level of setae in insertions), the discussion above on homology of tubercles observed in Neobelba applies as well.
Particularly interesting is the common presence of such subtle characters like the presence of 2 pairs of light spots on the notogaster near the insertions of l a and l p in most of Pateribelba species studied and in Neobelba ; or the form and size of setae on leg segments (e.g. dorsal setae on trochanters, femora and genua or length and form of solenidia except solenidion of tibia IV) very similar e.g. in both M.(P.) sphagni Strenzke, 1950 and N. pseudopapillipes . These common characters further support inclusion of Neobelba into Metabelba . As argued above, we consider the presence of companion seta on tibia IV to be a character at generic level, and therefore reject the synonymy with Pateribelba and support the original proposal of Subías (2004), i.e. to place Neobelba as a separate subgenus of the genus Metabelba , with close relation to subgenus Pateribelba.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |