
Phyllium ( Phyllium ) yapicum Cumming and Teemsma, 2018
|
publication ID |
https://doi.org/10.5281/zenodo.3708254 |
|
publication LSID |
lsid:zoobank.org:pub:21ACA19E-D965-4CC0-A8C4-76C9918094E4 |
|
DOI |
https://doi.org/10.5281/zenodo.3717204 |
|
persistent identifier |
https://treatment.plazi.org/id/F23087BA-AC30-FFD9-4681-A3AAFB924892 |
|
treatment provided by |
Felipe |
|
scientific name |
Phyllium ( Phyllium ) yapicum Cumming and Teemsma |
| status |
sp. nov. |
Phyllium ( Phyllium) yapicum Cumming and Teemsma View in CoL , new species
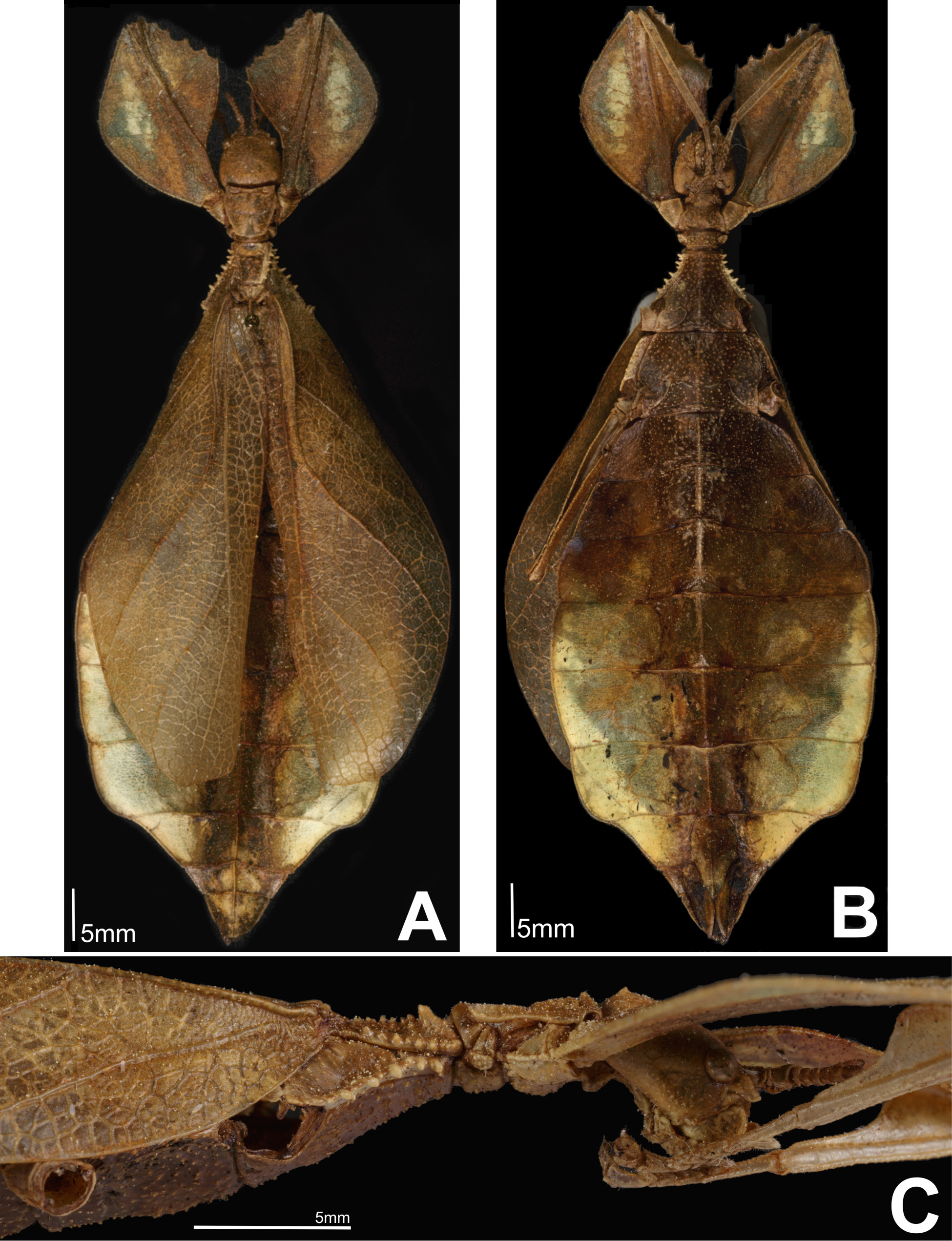
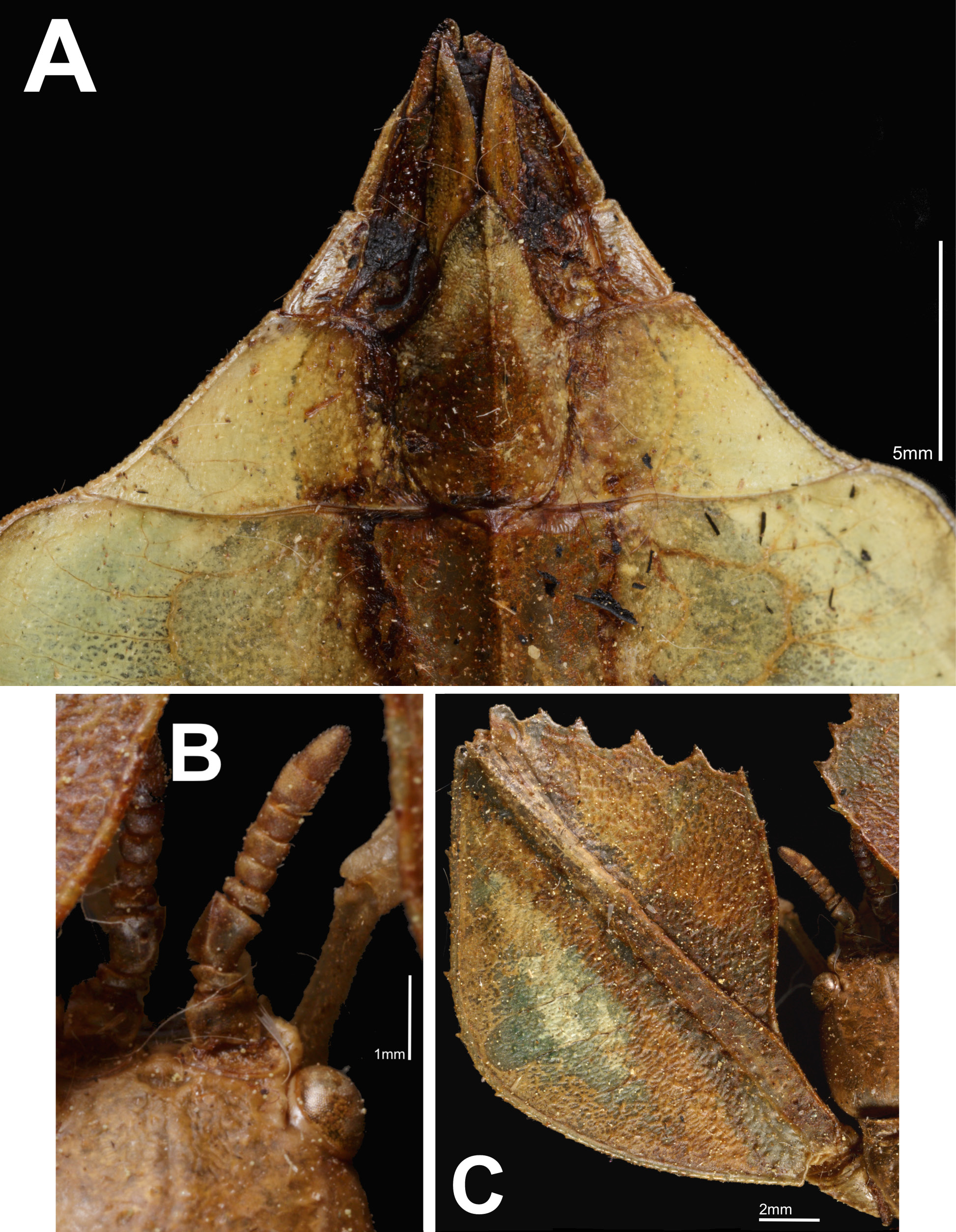
( Fig. 2A–C View Figure 2 , 3A–C View Figure 3 , 4A–C View Figure 4 )
Holotype. Female: FEDERATED STATES OF MICRONESIA, Yap Island, Kaday : XI-10-1980 (M. R. Lundgren, Collector): Deposited in the California Academy of Natural Sciences type collection, red type label: CASTYPE #19438.
Differentiation. The exterior lobe of the profemora is wider than the interior lobe and with developed alae, Phyllium ( Phyllium) yapicum n. sp. is placed within the celebicum species group, as described by Hennemann et al. 2009. Phyllium ( Ph.) yapicum n. sp. appears most similar in its morphology to Phyllium ( Ph.) ericoriai , Phyllium ( Ph.) bonifacioi , and Phyllium ( Ph.) tibetense because of the form of the thorax and the broad boxy shape of the abdomen. From all three species Phyllium ( Ph.) yapicum n. sp. can be differentiated by the unique interior lobe of the protibiae which is mostly situated in the proximal half; the distal half is reduced to a slender sliver of a lobe ( Fig. 4A View Figure 4 ).
Phyllium ( Ph.) yapicum n. sp. can further be differentiated from these species by several additional features. It can be differentiated from Phyllium ( Ph.) tibetense by the lack of lobes on the exterior pro- and metatibiae, since Phyllium ( Ph.) tibetense has clear but small exterior lobes on all tibiae. This is one of the few features that differentiate the two species as both species share many similar morphological characteristics such as mesopleurae that are prominently marked with tubercles ( Fig. 4B View Figure 4 ), an exterior lobe of the profemora with notable serration ( Fig. 3C View Figure 3 ), and an abdomen shape that is identical. The distant distribution of these two species, SW-China (Tibet) for Ph y llium ( Ph.) tibetense and Micronesia for Phyllium ( Ph.) yapicum n. sp., seems unlikely that they are of the closest relation despite their numerous morphological similarities.
A more likely relation to Phyllium ( Ph.) yapicum n. sp. would be the two celebicum species group members from the Philippines, Phyllium ( Ph.) ericoriai and Phyllium ( Ph.) bonifacioi . Both of these species have rather boxy abdominal shapes similar to that of Phyllium ( Ph.) yapicum n. sp., and they possess a thorax with mesopleurae that distinctly diverge throughout the entire length. From both species, Phyllium ( Ph.) yapicum n. sp. can be differentiated by the exterior lobe of the profemora with distinct serration ( Fig. 3C View Figure 3 ) while Phyllium ( Ph.) ericoriai and Phyllium ( Ph.) bonifacioi have smooth exterior lobes, and also by the mesopleurae with prominent tubercles ( Fig. 4B View Figure 4 ), as Phyllium ( Ph.) ericoriai and Phyllium ( Ph.) bonifacioi have mesopleurae with finer serration and lack of prominent tubercles.
Unfortunately, only the female is known for Phyllium ( Ph.) yapicum n. sp. lending an incomplete look into its likely relation. If the eggs and male morphology were known, it is likely a clearer position within the celebicum species group could be inferred. Eggs from both Phyllium ( Ph.) ericoriai and Phyllium ( Ph.) bonifacioi possess a rather unique morphology when compared to other phylliids. Therefore, it would have been ideal to compare eggs of Phyllium ( Ph.) yapicum n. sp. to these and other phylliids. However, due to the poor condition of the holotype, coupled with the thin appearance of the abdomen, it does not appear to have lived long enough as an adult to develop many, if any, eggs. Thus, the authors refrained from attempting to remove eggs from the abdomen for examination. With males of Phyllium ( Ph.) ericoriai and Phyllium ( Ph.) bonifacioi having rather different morphologies, Phyllium ( Ph.) ericoriai with a boxy abdomen and Phyllium ( Ph.) bonifacioi with a rounded abdomen, it will be interesting to discover the male morphological structure of Phyllium ( Ph.) yapicum n. sp. to see if it resembles either species.
Coloration. Overall coloration a pale yellow mixed with light brown, darker brown along the midline, thorax, and head likely from rot or being initially preserved in alcohol. Like other members of the family Phylliidae , it is assumed to be a vibrant green in life.
Morphology. Head capsule slightly longer than wide, the vertex is irregular with small nodes throughout and a slight sagittal furrow on the anterior half of the vertex. Posteromedial tubercle with a single point, no notable setae on or around the posteromedial tubercle. Frontal convexity broad at first but then quickly tapering to a rounded point and slightly bending upward at the apex, sparsely covered in thin transparent setae. Antennal fields similar in size to the compound eyes, not significantly broad. Anten- nal fields with the interior margin flush to the base of the scapus, lateral margin slightly wider than base of scapus. Frontal suture between antennal fields, small but distinct. Antennae consisting of nine segments, segments I–VII with few, sparsely spaced setae. Segments VIII and IX with dense, pale setae. Segment IV somewhat compressed when compared to the other terminal segments, similar in height to the pedicellus. Pars stridens on segment III with 36–40 fine teeth. Compound eyes moderately sized and slightly protruding away from the head capsule. Pronotum with an anterior margin that is slightly concave with a relatively smooth but distinct rim. Lateral margins of pronotum slightly diverge on the anterior third followed by straight converging margins to the posterior rim. Posterior rim slightly convex with a weak rim, width slightly more than half of the anterior rim width. Pronotum lateral and posterior with moderate rims which are marked by unevenly sized and spaced nodes. Face of the pronotum is marked by a distinct pit near the center and a slightly larger pit near the anterior rim along the sagittal plane, the remainder of the pronotum face is slightly textured with slight granulation. Prosternum is uniformly granulous, with nodes of even size and slightly uneven spacing. Meso- and metasternum heavily marked throughout with randomly spaced granules. Granulation of the metasterum continues onto abdominal sternite II which is fully covered in granulation, and partially onto sternites III and IV. Mesopraescutum anterior margin approximately as wide as long and lateral margins converge to a slightly narrower posterior. Anterior rim prominent and covered with irregularly granulose nodes in addition to two notable nodes at the apex. Lateral margins of the mesopraescutum with five to six major tubercles of uneven size, and three to four additional small nodes mixed throughout. Mesopraescutum surface irregularly granulose with those along the sagittal plane slightly more prominent and more densely packed. Mesopleurae starting at the anterior margin, uniformly diverging towards the posterior. Mesopleurae lateral margin with five major tubercles and five to six minor tubercles, which can be directly adjacent to, or evenly spaced between major tubercles. Face of the mesopleurae with a prominent pit on the anterior quarter, and a slight pit near the center of the mesopleurae, the remainder of the surface is slightly lumpy and marked with several small granules. Tegmina extending approximately half-way through abdominal segment VII. Alae moderately developed, extending into abdominal segment III. Abdominal segments II through the anterior half of IV uniformly diverging, posterior half of IV through the anterior two thirds of abdominal segment VII subparallel. The posterior one third of VII through the anal abdominal segment converge to a rounded apex. Subgenital plate stout, only slightly projecting under the anal abdominal segment and ending in a broad point ( Fig. 3A View Figure 3 ). Cerci only slightly cupped and with a lumpy irregular surface. Gonapophyses long and slender reaching the apex of the anal abdominal segment ( Fig. 3A View Figure 3 ). Profemora exterior lobe broad with an obtuse angle, wider than interior lobe. Profemora exterior lobe anterior half marked with four to five small serrate teeth pointing anteriorly. Profemora interior lobe posterior half smooth except for a small well-defined tooth on the anterior fifth. Anterior half with five large prominent looping teeth evenly spaced ( Fig. 3C View Figure 3 ). Mesofemora and mesotibia absent from holotype. Exterior lobe of metafemora thin and lacking teeth, relatively straight and hugging the shaft of the metafemora. Interior lobe of the metafemora with seven serrate teeth on the distal half. Protibiae lacking exterior lobe, interior lobe only on the proximal two thirds, in the shape of a scalene triangle. Metatibiae simple, lacking lobes.
Measurements of holotype [mm]. Length of body (including cerci and head, excluding antennae) 73.4, length/greatest width of head 6.3/5.9, pronotum 4.7, mesonotum 6.9, length of tegmina 44.5, alae length approximately 12.0-13.0 (accurate measurement unavailable due to tegmina obscuring view), greatest width of abdomen 28.9, profemora 17.9, mesofemora missing from specimen, metafemora 17.2, protibiae 12.1, mesotibiae missing from specimen, metatibiae 13.2, antennae 3.9.
Distribution. Currently only known from the type locality, Kaday, Yap Island, Micronesia. With the Phylliidae being such cryptic insects, it is not surprising that they have gone overlooked on this small island nation. It would be interesting to know if this species or congenerics also occur on the other nearby islands. The country of The Republic of Palau, with larger islands than Yap Island, and with a closer proximity to other phylliid rich areas, may eventually reveal additional species. Even the island of Guam could be a possibility since it is situated to the northeast of Yap Island. It is hoped further examination of material collected from Micronesia and nearby islands will reveal the geographic distribution with more clarity.
Etymology. This species is named for the type locality: Yap Island, Micronesia. As a significant range expansion for the family, it is fitting to use an adjectival name for this notable occasion.

| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |