
Pseudalataspora lophii, Afonso-Dias, Isabel, Kalavati, Chaganti, Mackenzie, Ken & Mackenzie, Kevin S., 2007
|
publication ID |
https://doi.org/10.5281/zenodo.176573 |
|
DOI |
https://doi.org/10.5281/zenodo.5625784 |
|
persistent identifier |
https://treatment.plazi.org/id/039887D5-7C07-FFFE-FDCC-AFCE64C3CFC3 |
|
treatment provided by |
Plazi |
|
scientific name |
Pseudalataspora lophii |
| status |
sp. nov. |
Pseudalataspora lophii n. sp. ( Figures 10–15 View FIGURES 10 – 15 )
Material studied
Host: Lophius piscatorius L.
Site of infection: gall bladder
Locality, date and depth: (1) 58°51ˏN, 07°35ˏW (NW of Outer Hebrides), 30 March, 2000, 250m; (2) 58°45ˏN, 07°49ˏW (NW of Outer Hebrides), 30 March, 2000, 400m; (3) 58°29ˏN, 08°54ˏW (NW of Outer Hebrides), 30 March, 2000, 470m; (4) 58°45ˏN, 08°03ˏW (NW of Outer Hebrides), 30 March, 2000, 700m; (5) 57°20ˏN, 05°50ˏW (Off Skye, Inner Hebrides), March 2004, depth unknown; (6) 57°20ˏN, 05°50ˏW (Off Skye, Inner Hebrides), August, 2004, depth unknown.
Prevalence: (1) 3 of 16 (19%); (2) 14 of 26 (54%); (3) 1 of 1; (4) 1 of 1; (5) 1 of 2; (6) 1 of 2.
Double infections: two host individuals, one each from positions (1) and (2), had double infections of P. lophii and C. lophii .
Host length range: (1) 41–64 cm; (2) 45–66 cm; (3) and (4) lengths unknown.
Collection number: 2005:7:1:3.
Description
Trophozoite ( Figures 10, 11 View FIGURES 10 – 15 ) disporous, ovoid with a single lobopodium sometimes bifurcating at its tip, which is attached to the wall of the gall bladder. Ectoplasm dense and clearly demarcated. Endoplasm fine, lightly staining. Dimensions, based on 8 fixed specimens: 25.6–35.2 x 14.4–32.0.
Spore ( Figures 12, 13, 14, 15 View FIGURES 10 – 15 ,) triangular or rhomboidal with pointed tips in sutural view, flat and curved in valvular view. Sutural line prominent and raised (this feature can be seen in Figure 14 View FIGURES 10 – 15 , but is obscured in Figures 12 and 13 View FIGURES 10 – 15 by the enveloping alate processes). Sporoplasm binucleate. Valve shells drawn out into two delicate broad alate processes that join together at their proximal extremities forming a parachute-like structure over the valves. The valves are often observed folded over with the alate processes covering them ( Figure 13 View FIGURES 10 – 15 ). Polar capsules oval, terminal, in close proximity to one another and overlapping in sutural view. Polar filament wide, flat, deeply staining and with only 1.5–2 coils. Dimensions, based on 30 fixed spores, as ranges with means ± SD in parentheses: spore length 7.2–10.0 (9.8 ± 1.6); spore width, including alate processes 21.6–28.8 (25.9 ± 2.8); spore width, excluding alate processes 16.8–22.6 (18.2 ± 1.2); spore thickness 15.2 – 20.8 (17.5 ± 1.01); polar capsule length: 3.8–5.4 (4.3 ± 0.7); polar capsule width: 3.0–4.0 (3.6 ± 0.2); polar capsule length: spore length = 1: 2.0–3.2; spore length: spore width = 1: 3.2–4.3.
Discussion
Pseudalataspora lophii n. sp. differs from the other 11 species of Pseudalataspora described to date in the dimensions of the spore ( Table 3 View TABLE 3. A ) and in having alate processes that cover the spore body. The numbers of coils of the polar filament are not given in the descriptions of most of these species, but the figures indicate that none has less than three, as compared with a maximum of only two coils for P. lophii n. sp.
The function of the parachute-like alate processes that cover the spore of P. lophii would appear to be to facilitate the distribution of the spore after release from the host. Lophius spp. are ambush predators that immerse themselves in the substrate with a shuffling motion (Laurensen et al., 2004). This behaviour would tend to throw myxosporean spores up and away from the infected host and the alate processes would then assist in their further dispersal. Whether this might result in direct infection of other anglerfishes or of an invertebrate alternative host is unknown. Only one marine myxosporean life cycle has so far been shown to require an invertebrate host ( Køie et al., 2004), although actinosporean stages have been found in several species of marine polychaetes and oligochaetes. On the other hand, there is also evidence of direct fish-to-fish transmission of marine myxosporeans ( Diamant, 1997; Yasuda et al., 2002).
As with the genus Alataspora , Pseudalataspora has been referred to erroneously in the literature as Pseudalatospora or Pseudoalatospora , the latter being the misspelling by Lom & Dykova (1992).
Three species of myxosporean are now known to infect the gall bladders of L. piscatorius , while two species are known to infect the gall bladders of L. budegassa . Evidence from the literature and from the present study suggest that these parasites have different endemic areas, which makes them potentially useful candidates as biological tags in population studies of their fish hosts. Recent genetic studies have suggested that stocks of L. piscatorius in the northern part of its distribution are part of a single more or less panmictic reproductive population ( O’Sullivan et al., 2006). Parasites, however, can often be used to identify subpopulations of fish distinguished by certain behavioural differences, but which genetic studies may not pick up because there is still a considerable amount of gene flow between them ( MacKenzie & Abaunza, 2005). In future studies we plan to examine more anglerfish samples from different parts of their geographical distribution to test if their myxosporean gall bladder parasites can be used to reveal finer details of host population structure.
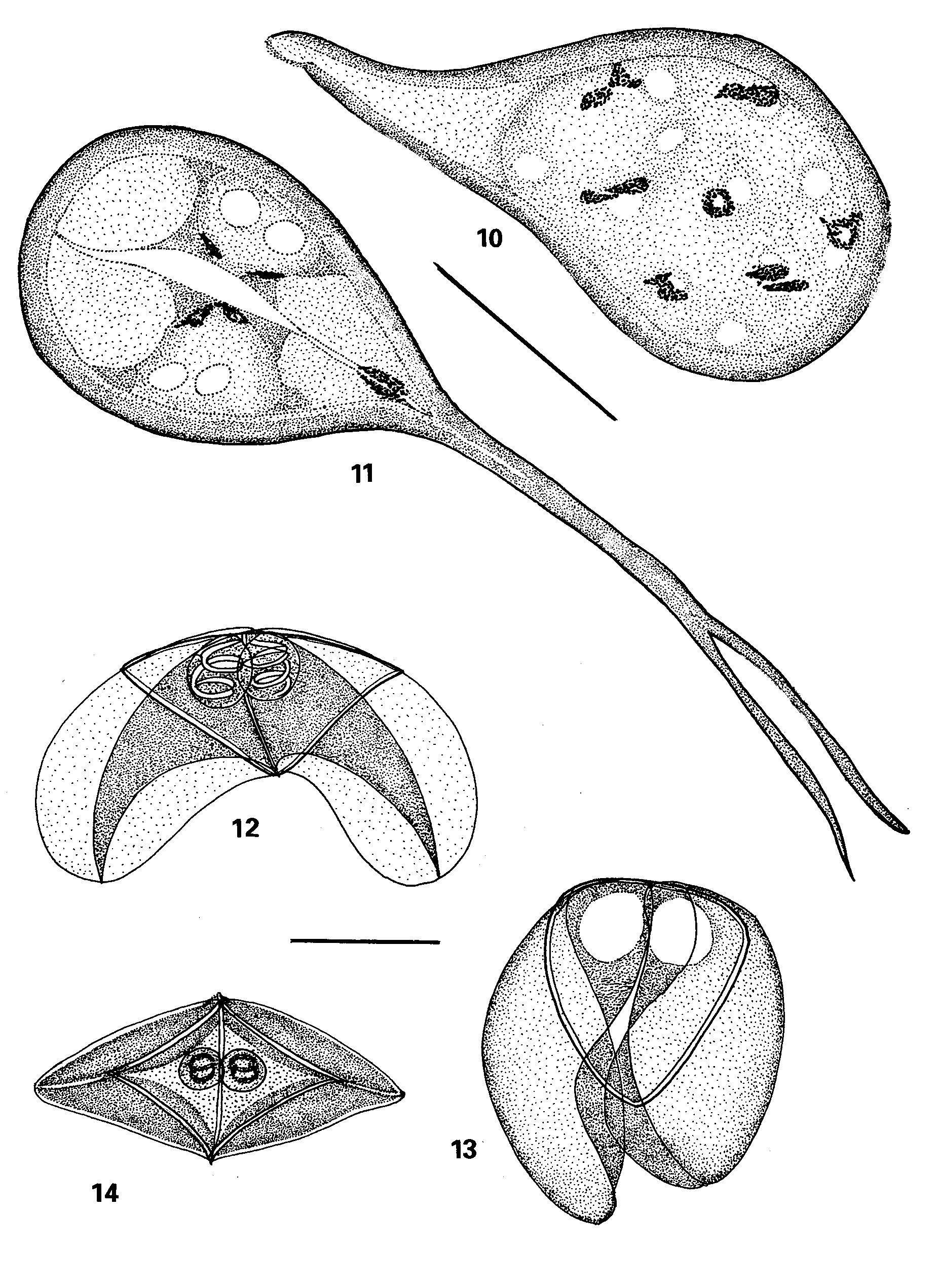
TABLE 3. A comparison of the spore dimensions of Pseudalataspora lophii with those of the other species in the genus. Measurements are in micrometres. ND = no data.
| Species of Pseudalataspora P. lophii n. sp. P. atlantica Kovaleva, Velev | Spore Length Thickness 7.2–10.0 21.6–28.8 6.7–10.0 34.6–42.5 | Polar capsule Length Width 3.8–5.4 3.0–4.0 3.3–4.0 3.3–4.0 | Host Lophius piscatorius Chlorophthalmus agassizi , | Locality Off Northwest Scotland Atlantic Ocean off | ||
|---|---|---|---|---|---|---|
| & Vladev, 1993 | C. atlanticus , Caranx rhon- | Africa | ||||
| P. beryxi Kovaleva & Gae- | 5.3–6.65 14.6–19.95 | 1.5 1.5 | chus, Fistularia tabacaria Beryx splendens | Atlantic Ocean | ||
| vskaya, 1988 P. indecora Kovaleva, Velev | 3.3–4.7 13.4–20.0 | 2.0 2.0 | Dentex angolensis | Atlantic Ocean off | ||
| & Vladev, 1993 P. insolita Kovaleva, Velev & | 5.3–6.0 14.6–17.3 | 2.0 2.0 | Caranx rhonchus | Africa Atlantic Ocean off | ||
| Vladev, 1993 P. meridionalis Kovaleva , | 6.7–9.5 16–20 | 2.5–2.7 2.5–2.7 | Notothenia rossii | Africa Antarctic Ocean | ||
| Rodjuk & Grudnev, 2002 P. originalis Aseeva, 2003 | 10–12 27–36 | 3.5–4.0 2.8–3.0 | Hippoglossoides dubius , | Sea of Japan | ||
| P. pontica Kovaleva, Donets | 5.5–6.5 16.0–19.9 | 3.0 2.7 | Glyptocephalus stelleri Liza aurata | Black Sea | ||
| & Kolesnikova, 1989 P. s c o m b r i Kovaleva & Gae- | 4.65–5.98 7.92–10.64 | 1.3–1.5 1.3–1.5 | Scomber japonicus | Southeast Pacific Ocean | ||
| vskaya, 1983 P. sebastei Bakay & Grudnev , | 13.3 22.64 | 4.65 4.65 | Sebastes marinus , S. men- | North Atlantic | ||
| 1998 | vivipaP. squamifrons Kovaleva, | 5.3–6.7 19.9–21.3 | 2–2.3 1.3 | tella, S. fasciatus , S. vivipaP. squamifrons Kovaleva, 5.3–6.7 19.9–21.3 2–2.3 1.3 rus Lepidonotothen squamifrons Antarctic Ocean | rus Lepidonotothen squamifrons | Antarctic Ocean |
| vivipaP. squamifrons Kovaleva, | 5.3–6.7 19.9–21.3 | 2–2.3 1.3 | rus Lepidonotothen squamifrons | Antarctic Ocean | ||
| Rodjuk & Grudnev, 2002 P. umbraculiformis Gae- | 8.0–9.3 14.6–17.3 | 2.7 2.7 | Gaidropsarus mediterraneus | Celtic Sea | ||
| vskaya & Kovaleva, 1984 | ||||||
| General Discussion |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Myxosporea |
|
Order |
|
|
Family |
|
|
Genus |